













INTRODUCTION
Milk is consumed for human health, nutrition, and immune function because of its essential nutrients and natural bioactive components [1]. Global milk production is forecast to steadily increase from 887 metric tons in 2021 to 1,060 metric tons in 2031 [2]. However, the global population of 8.0 billion in 2022 is projected to increase to 8.7 billion by 2032 and 10 billion by 2050 [3–5]. While the current population of cows utilized for milk production stands at nearly 234 million [6], the demand for milk production is projected to persistently rise.
Over the past 70 years, the dairy industry has been guided by dairy research and policies aimed at enhancing the economic efficiency of milk production [7]. Consequently, dairy cows in current dairy farming have been raised to increase milk production in concentrated animal-feeding operations [8]. However, the dairy farming has negatively impacted the environment and animal welfare [7]. To mitigate the negative environmental impacts of dairy farming, a Dairy Sustainability Framework has been established by the global dairy sector [9]. Also, in alignment with sustainable dairy farming practices, several researchers in the field are dedicated to developing cow milk proteins through precision fermentation technology and cell culture. [10,11].
The milk proteins and fats are synthesized within the epithelial cells (ECs) of the mammary gland [12] The mammary gland is comprised of luminal epithelium and basal epithelium. The luminal epithelium consists of ECs, while the basal epithelium consists of fibroblasts (FBs), myoepithelial cells (MCs), and adipocytes [13]. According to a previous study, milk productivity is directly related to the number of mammary ECs [14]. This indicates that the establishment of a bovine mammary ECs is essential for the production of milk proteins and fats in in vitro cell culture systems. However, in fact, primary bovine mammary ECs exhibit cellular instability because of their finite lifespan [15,16].
Previously, primary bovine mammary EC models have been established to study the synthesis of milk components and mammary gland function [17–19]. Most of these studies have evaluated cellular characteristics at early passages via the analysis of morphology, cell-specific markers, growth patterns, and secretion of milk components. Nevertheless, the development of a reliable primary bovine mammary alveolar cell model remains a challenge, especially as passages progress. A prior study suggested that enhancing cellular stability can be achieved by assessing cellular characteristics during cell growth and bioproduction across successive passages. [20]. Hence, it is important to conduct a comprehensive assessment of cellular characteristics and milk component productivity across successive passages to ensure the stability of primary bovine mammary ECs.
Overall, the cellular characteristics of the primary bovine mammary ECs during serial passage have not been comprehensively studied, mainly due to cellular instability. Therefore, this study aimed to assess the cellular characteristics and milk component productivity of primary bovine mammary ECs throughout cell subculturing, with the goal of identifying areas for enhancing the stability of cells responsible for the sustainable production of cell-cultured milk.
MATERIALS AND METHODS
Dulbecco’s modified Eagle medium/nutrient mixture F12 (DMEM/F12) was obtained from Gibco (Grand Island, NY, USA). Fetal bovine serum (FBS), penicillin/streptomycin (P/S), and trypsin-ethylenediaminetetraacetic acid (EDTA) were supplied by WELGENE (Gyeongsan, Daegu, Korea). Gentamicin sulfate was obtained from Sigma-Aldrich (St. Louis, MO, USA). Amphotericin B was purchased from Gibco. Insulin (INS), cortisol (CORT), progesterone (P4), 17-β estradiol (E2), and epidermal growth factor (EGF) were purchased from Sigma-Aldrich. Prolactin (PRL) was obtained from ProSpec (Ness-Ziona, Israel). Phosphate-buffered saline (PBS) was purchased from Lonza (Walkersville, MD, USA). Hank’s balanced salt solution (HBSS) was provided by Gibco. The control culture media (CCM) were freshly prepared with DMEM/F12 containing 10% FBS, 1% P/S, 100 μg/mL gentamycin, and 5 μg/mL amphotericin B. The differentiation culture media (DCM) were prepared with CCM added with 1 μg/mL PRL, 5 μg/mL INS, 1 μg/mL CORT, 5 μg/mL P4, 5 μg/mL E2, and 10 ng/mL EGF.
Primary bovine mammary cells were isolated from the bovine mammary parenchymal tissues of one lactating Holstein dairy cows (53-month-old and 306 kg body weight) after slaughter. Fresh pieces of the mammary tissue were placed in the sterilized specimen cups with HBSS supplemented with penicillin (200 U/mL), streptomycin (200 μg/mL), gentamycin (200 μg/mL), and amphotericin B (10 μg/mL) and immediately transported to the laboratory. Five grams of mammary tissue was cut into 1 cm3 pieces, centrifuged at 1,500×g for 5 min, and washed three times with HBSS to remove blood and milk. The rinsed pieces of tissues were transferred onto a 10-cm cell culture dish using 1 mL sterile tips (SPL, Pocheon, Korea) and were carefully incubated at 37° in a humidified 5% CO2 incubator using CCM. The CCM was replaced every 2 d until the cells grow out from the tissues to the bottom of the 10-cm cell culture dish. After the removal of mammary tissues, FBs, MCs, and ECs were isolated and purified using trypsin-EDTA solution based on their different trypsin sensitivity of 2, 2, and 10 min, respectively. ECs were differentiated using DCM for 5 d, and DCM was replaced every 2 d. The morphology of the cells was imaged and captured using Nikon Eclipse Ti2-U and Nikon Eclipse Ts2R cameras (Nikon, Tokyo, Japan).
Cytoskeletal protein levels were determined by immunofluorescence staining. FBs, MCs, and ECs were cultured with CCM at a density of 0.05 × 106 cells per well in 12-well cell culture plates for 7 d and fixed with 4% paraformaldehyde for 15 min. Then, cells were treated with 0.1% Triton X-100 in PBS for permeabilization for 10 min. The cells were then blocked using 3% bovine serum albumin for 90 min and incubated with anti-mouse cytokeratin 18 (CK18; 1:200, Santa Cruz Biotechnology, Dallas, TX, USA), anti-mouse CK19 (1:200, Progen Biotechnik GMBH, Heidelberg, Germany), anti-rabbit vimentin (1:500, Thermo Fisher Scientific, Rockford, IL, USA), and anti-rabbit α-smooth muscle actin (α-SMA; 1:500, Thermo Fisher Scientific) diluted in 3% bovine serum albumin at 4°C for 15 h. Subsequently, the cells were washed three times with 0.1% tween 20 in PBS and incubated with DyLight 488 conjugated-donkey anti-rabbit IgG H&L (1:1,000, Bethyl Laboratories, Montgomery, TX, USA) and goat anti-mouse IgG H&L FITC (1:1,000, Abcam, Cambridge, UK) at 25°C for 90 min. After washing three times with 0.1% tween 20 in PBS, the cells were fixed with 4% paraformaldehyde. The nuclei were stained with 4,6-diamidino-2-phenylindole dihydrochloride (DAPI; 1 µg/mL) for 10 min and washed three times with 0.1% tween 20 in PBS. The stained cells were imaged and captured using a Nikon Eclipse Ti2-U and Nikon Eclipse Ts2R camera.
FBs, MCs, and ECs were grown with CCM for 7 d at a density of 0.1 × 106 cells per well in 6-well cell culture plates. ECs were cultured with CCM and DCM for 5 d at a density of 0.1 × 106 cells per well in 6-well cell culture plates. Radioimmunoprecipitation assay buffer (Elpis Biotech, Daejeon, Korea) supplemented with a protease inhibitor cocktail (Abbkine, Atlanta, GA, USA) was used to lyse the cells. The cell lysates were collected in a 1.7 mL microtube and centrifuged at 17,000 ×g at 4°C for 20 min. Proteins were quantified using a bicinchoninic acid protein assay kit (Thermo Fisher Scientific). Protein samples were loaded into sample wells of stacking (5%) and separating (10%) acrylamide gels and separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis. The loaded proteins were transferred onto nitrocellulose membranes (GE Healthcare Biosciences, Pittsburgh, PA, USA) using a Semi-Dry Electrophoretic Transfer Cell (Bio-Rad Laboratories, Hercules, CA, USA). The nitrocellulose membrane was blocked with 5% nonfat milk buffer dissolved in Tris-buffered saline with Tween 20 buffer at 25°C for 90 min and incubated with anti-mouse CK 18 (1:500), anti-mouse CK19 (1:500), anti-rabbit vimentin (1:3,000), anti-rabbit α-SMA (1:3,000), anti-rabbit E-cadherin (1:20,000, Merck Millipore, Darmstadt, Germany), anti-rabbit occludin (1:3000, Invitrogen), anti-rabbit claudin-1 (1:3,000, Merck Millipore), anti-rabbit α-tubulin (1:5,000, Cell Signaling Technology, Danvers, MA, USA), and anti-glyceraldehyde 3-phosphate dehydrogenase (GAPDH; Merck Millipore) at 4°C for 1–3 d. After washing three times with PBS for 15 min, the membranes were incubated with goat anti-rabbit IgG conjugated with horseradish peroxidase (Enzo Life Sciences, Lausen, Switzerland) and goat anti-mouse IgG H&L conjugated with horseradish peroxidase (Abcam). The signals of proteins were visualized using enhanced chemiluminescence detection reagents (Thermo Fisher Scientific). α-Tubulin and GAPDH were used as internal controls for normalization. The intensities of protein bands were quantified using Image J software (National Institute of Health, Bethesda, MD, USA).
The growth characteristics of FBs, MCs, and ECs cultured with CCM and DCM were evaluated using trypan blue solution (Gibco). The cells were seeded at a density of 0.05 × 106 cells per well in a 12-well cell culture. The cells were dissociated using trypsin-EDTA solution for 1, 3, 5, or 7 d. After staining the cells with trypan blue solution, viable cells were counted using a hemocytometer (Hausser Scientific, Horsham, PA, USA).
Gene expression levels related to milk proteins (CSN1S1, CSN2, CSN3, ALA, and BLG) were evaluated using real-time polymerase chain reaction (RT-PCR). ECs were cultured and differentiated with CCM and DCM at a density of 0.05 × 106 cells per well in 12-well cell culture plates for 5 d. Total RNA was extracted using TRIzol (Thermo Fisher Scientific, Carlsbad, CA, USA) and cDNA was synthesized using the TOPscript RT DryMIX kit (Enzynomics, Daejeon, Korea). The mRNA expression levels were analyzed using 2× RT-PCR Smart mix (BIOFACT, Daejeon, Korea) and an RT-PCR system (Roche LightCyclerⓇ 96 System, Basel, Switzerland) with thermal cycling conditions of 95°C for 15 min and 60 cycles of denaturation at 95°C for 10 s, annealing at 60°C for 10 s, and extension at 72°C for 10 s. The mRNA expression levels were quantified using the 2−ΔΔCt method and GAPDH was used as an internal control for normalization. The primer sequence (BIONICS, Seoul, Korea) designed using the AmplifX software version 1.7.0 as follows: CSN1S1, (F) 5’- ACT GAG GAT CAA GCC ATG GAA G-3, (R) 5’-GAA TGT GCT TCT GCT CAA CAC T-3’; CSN2, (F) 5’-CTG GAA TTA ACT GCT TCT ACC T-3, ’ (R) 5’-TAC TCT GCG ATT TGT CTT ATT GA-3’; CSN3, (F) 5’-GGC GAG CCT ACA AGT ACA CCT A-3, ’ (R) 5’-GGA CTG TGT TGA TCT CAG GTG G-3’; ALA, (F) 5’-CCT GAA TGG GTC TGT ACC ACG TTT-3, ’ (R) 5’-ATG TTG CTT GAG TGA GGG TTC TGG-3’; BLG, (F) 5’-AGG CCT CCT ATT GTC CTC GT-3, ’ (R) 5’-GCA AAG GAC ACA GGG AGA AG-3. ’ GAPDH (F) 5’-ATG ATT CCA CCC ACG GCA AGT T-3,’ and (R) 5’-ATC ACC CCA CTT GAT GTT GGC A-3.’
Oil red O dye (Sigma-Aldrich) was used to evaluate the levels of intracellular triglycerides, major components of milk fat. ECs were cultured with CCM and DCM at a density of 0.05 × 106 cells per well in 12-well cell culture plates for 5 d. The cells were fixed in 10% formalin at 25°C for 1 h. After fixation, cells were washed with 60% isopropanol and allowed to dry completely. Subsequently, the cells were stained with the oil red O working solution for 10 min. After washing three times with deionized distilled water to remove the unbound dye, the stained cells were imaged and captured using Nikon Eclipse Ti2-U and Nikon Eclipse Ts2R cameras. The intensities of the oil red O-stained areas were quantified using ImageJ software.
α-Casein and triglycerides contents in culture media were analyzed using Bovine Casein Alpha (CSN1) enzyme-linked immunosorbent assay (ELISA) Kit (Bioss Antibodies, Woburn, MA, USA) and Triglyceride Assay Kit (Abcam). In brief, ECs were cultured with CCM and DCM at a density of 0.05 × 106 cells per well in a 12-well cell culture plate for 5 d. After that, CCM and DCM were collected in 1.7 mL micro tubes and centrifuged at 17,000×g at 4°C for 20 min. The supernatants of CCM and DCM were preserved at –80°C until use. The ELISA was performed according to the instructions of the manufacturers.
The matrix metalloproteinase (MMP) activity was determined as described previously, with slight modifications [21]. ECs were cultured with CCM and DCM at a density of 0.05 × 106 cells per well in 12-well cell culture plates for 5 d. Subsequently, CCM and DCM were collected in 1.7 mL micro tubes and centrifuged at 17,000×g at 4°C for 20 min. Proteins in the supernatants were quantified using a bicinchoninic acid protein assay kit. The CCM and DCM were analyzed using 8% sodium dodecyl sulfate-polyacrylamide gel electrophoresis containing 0.1% gelatin as the MMP substrate. After that, the gel was washed twice for 30 min with a washing buffer containing 12.5% Triton™ X-100. The gel was then incubated with reaction buffer solution supplemented with Tris-HCl of 40 mM, calcium chloride of 5 mM, and sodium azide of 3 mM at 37°C for 15 h. The activities of MMP-2 and -9 were determined by negative staining with Coomassie Brilliant Blue R 250 (Sigma-Aldrich).
All experiment were conducted at least three times independently. The experimental data were presented as mean ± SEM. Statistical significance was analyzed by Student’s t-test using SPSS-PASW statistics software (version 22.0, SPSS, Chicago, IL, USA). Statistical differences were considered significant at p < 0.05.
RESULTS
The parenchymal tissues of bovine mammary gland were dissected from the bovine udder (Fig. 1A). The 1 cm3 pieces of bovine mammary parenchymal tissues were placed on the bottom of 10-cm cell culture dish for inducing the growth of bovine mammary cells. After 12 d of tissue incubation in CCM, FBs were firstly isolated from the mammary parenchymal tissue (Fig. 1B). MCs and ECs were isolated from mammary tissue after 24 d of tissue incubation, where dome-like structure was observed. In addition, ECs were enveloped by MCs with the outer layer consisting of FBs (Fig. 1B). The isolated FBs, MCs, and ECs were purified using different trypsin digestion time, i.e., 2 and 10 min, respectively.

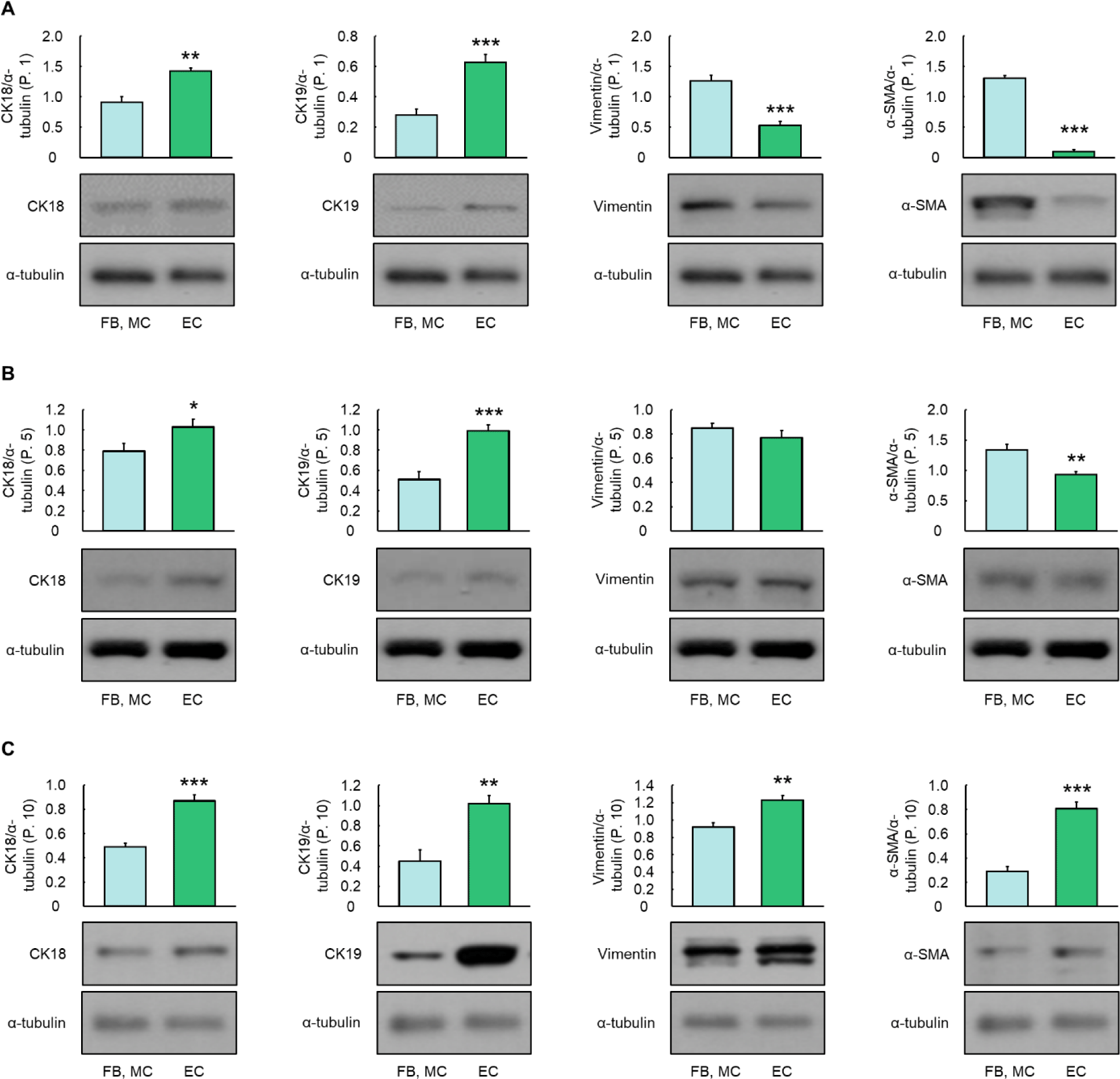
The fluorescence expression levels of CK18 and CK19 were markedly higher in ECs at passage 1 than in FBs and MCs (Fig. 2A). In contrast, the vimentin and α-SMA in FBs and MCs at passage 1 had higher fluorescence expression levels than ECs (Fig. 2A). CK18 and CK19 in ECs at passage 5 showed higher fluorescence expression levels than those in FBs and MCs. FBs and MCs at passage 5 had relatively higher fluorescence expression levels of α-SMA than ECs (Fig. 2B). However, there were no differences in the expression levels of vimentin among FBs, MCs, and ECs at passage 5 (Fig. 2B). At passage 10, fluorescence expression levels of CK18, CK19, vimentin, and α-SMA were slightly higher in ECs than in FBs and MCs (Fig. 2C).

At passage 1, CK18 and CK19 showed significantly higher protein expression levels than in FBs and MCs (Fig. 3A, p < 0.01). Also at passage 1, vimentin and α-SMA in FBs and MCs had significantly higher protein expression levels than ECs (Fig. 3A, p < 0.001). At passage 5, the protein expressions of CK18 and CK19 showed significant increases in ECs compared to FBs and MCs (p < 0.05, Fig. 3B). In contrast, the protein expression of α-SMA showed a significant increase in FBs and MCs at passage 5 compared to ECs (p < 0.01, Fig. 3B) and there was no significant difference among FBs, MCs, and ECs at passage 5 in the protein expression level of vimentin (p > 0.05, Fig. 3B). All protein expressions of CK18, CK19, vimentin, and α-SMA were significantly higher in ECs at passage 10 than FBs and MCs (p < 0.01, Fig. 3C).
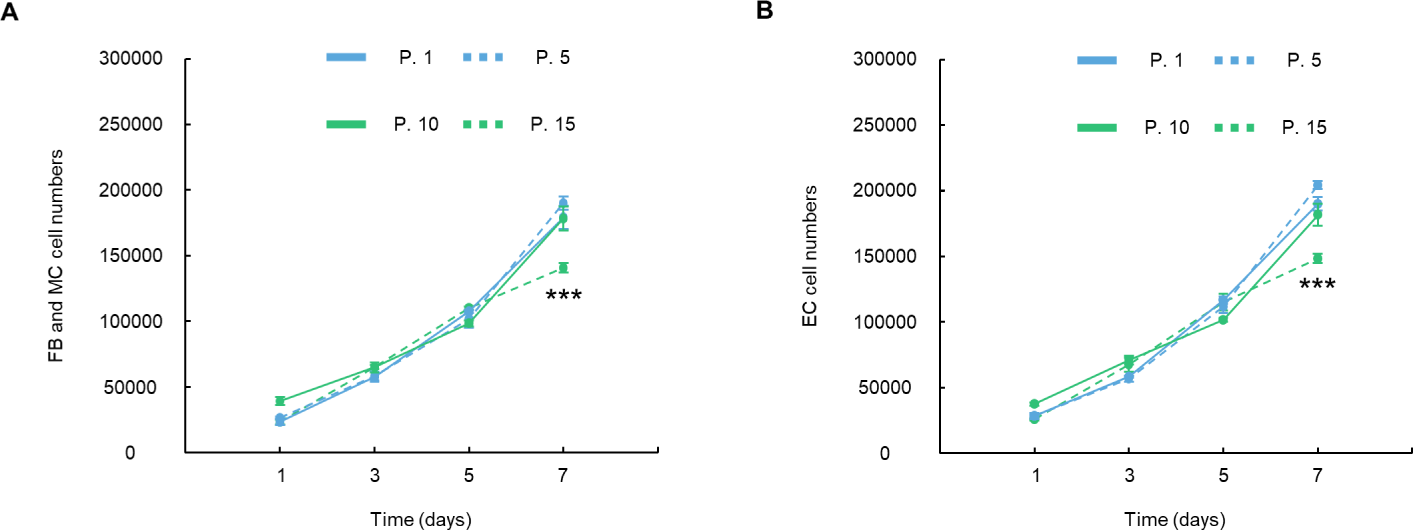
At passage 1, FBs and MCs grew from 2.33 × 104 at 1 d to 1.80 × 105 at 7 d (Fig. 4A). In contrast, ECs had cell numbers of 2.83 × 104 at 1 d and 1.90 × 105 at 7 d (Fig. 4B). However, at passage 15, the growth of FBs and MCs significantly decreased from 2.33 × 104 at 1 d to 1.41 × 105 at 7 d compared to that at passage 1 (p < 0.001, Fig. 4A). The growth of ECs significantly decreased from 2.58 × 104 at 1 d to 1.48 × 105 at 7 d (p < 0.001, Fig. 4B).
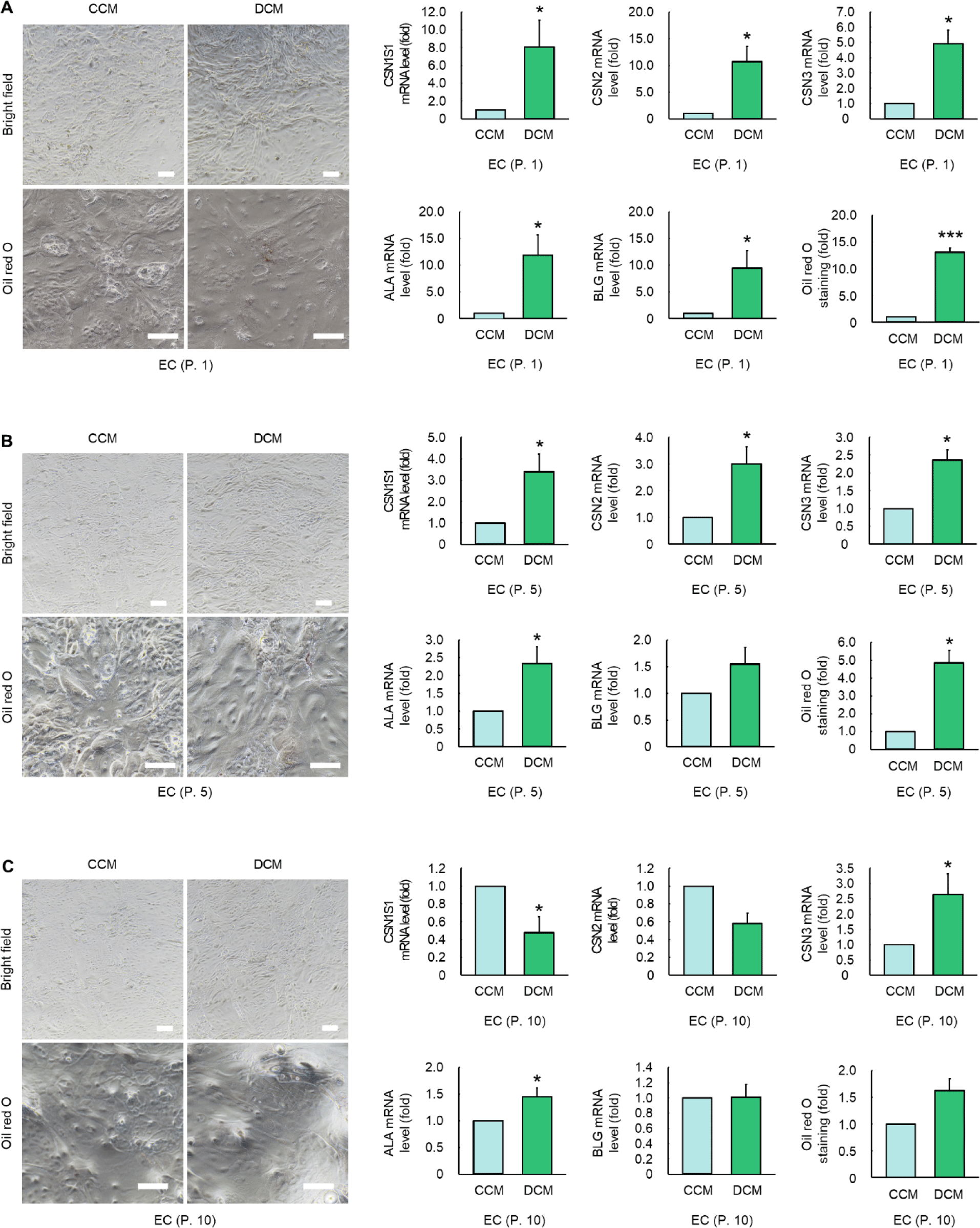
Culturing ECs with DCM for 5 d resulted in significant morphological changes compared with cells cultured with CCM (Figs. 5A, 5B, and 5C). Alveoli-like structural forms were observed in ECs cultured in DCM. However, a decrease in alveoli-like structures and an increase in vacuoles in ECs were observed in accordance with an increase in passage number (Figs. 5A, 5B, and 5C). At passage 1, ECs cultured in DCM exhibited significantly elevated expression levels (8.08-, 10.71-, 4.92-, 11.82-, 9.47-, and 13.12-fold) of milk protein-related genes (CSN1S1, CSN2, CSN3, ALA, and BLG) and displayed increased oil red O staining compared to those cultured in CCM (p < 0.05, Fig. 5A). At passage 5, the levels of milk protein-related genes and oil red O staining in ECs cultured with DCM were elevated to 3.39-, 4.757-, 2.639-, 2.621-, 1.55-, and 4.86-fold compared to the CCM (p < 0.05, Fig. 5B). However, at passage 10, ECs cultured in DCM did not show significant increases in the expression of milk protein-related genes or oil red O staining compared to those cultured in CCM (p > 0.05, Fig. 5C).
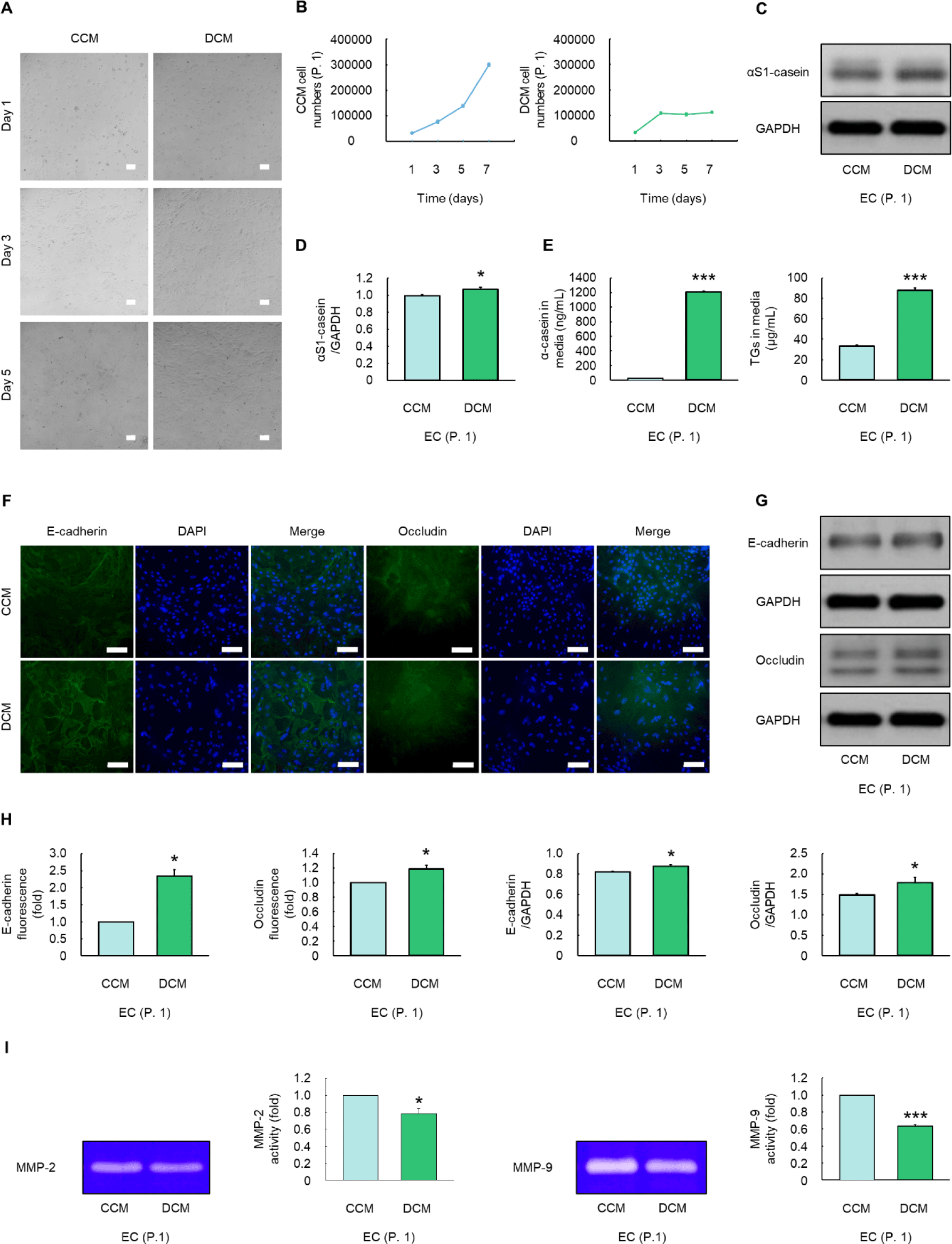
The CCM increased the number of ECs, whereas the DCM maintained the cell count (Figs. 6A and 6B). CCM exhibited morphological characteristics resembling those of vacuoles, whereas DCM displayed an alveolar structure (Fig. 6A). Moreover, ECs cultured in DCM significantly elevated the protein expression level of αS1-casein compared to those cultured in CCM (p < 0.05, Figs. 6C and 6D). ECs cultured with DCM had significantly higher α-casein and triglyceride levels in media than those cultured with CCM (p < 0.001, Fig. 6E). The green fluorescence and protein expression levels of tight junctions (TJs) including E-cadherin and occludin were significantly enhanced in ECs cultured with DCM than those cultured with CCM (p < 0.05, Figs. 6F, 6G, and 6H). The activity of MMP-2 and -9 in cells cultured in DCM was significantly lower than those in CCM (p < 0.05, Fig. 6I).
DISCUSSION
Interest in sustainable dairy farming is continuously increasing because of concerns about greenhouse gas emissions, nitrogen and phosphorus excretion from manure, and animal welfare resulting from intensive dairy farming [22]. In recent years, companies have used fermentation-based cellular agriculture to manufacture milk components like CSN and BLG [10]. However, this innovative precision fermentation technology has raised public concerns, particularly in relation to the utilization of genetically modified organisms [23]. Cellular agriculture in the dairy sector is driven by the goal of utilizing animal cells, specifically ECs derived from the alveoli of the mammary gland [24,25]. These cells have the capacity to generate essential milk components, including milk proteins and fats through secretory differentiation. Consequently, our research has focused on examining the cellular properties of primary bovine mammary cells and establishing an EC model for the production of milk constituents.
Mammary alveoli are fundamental components of the mammary glands that are responsible for milk production and secretion [24]. The parenchyma of alveoli is composed of inner milk secretory ECs that surround the lumen, outer MCs that attach to the base of the mammary epithelium, and the basement membrane that contacts the MCs. In addition, the stromal compartment comprises various stromal cells, such as FBs, adipocytes, endothelial cells, and the extracellular matrix [26]. Therefore, from the perspective of cellular structure, FBs and MCs are located on the outer side and ECs are positioned on the inner side of the mammary alveolus. In accordance with the structure of the alveolus, our results showed that FBs and MCs located on the outer side of the alveolus were first isolated after 12 d of tissue incubation, whereas ECs located on the inner side were isolated after 24 d (Fig. 1). A previous study reported that FBs and ECs were elongated after culturing for 5 and 10 d, respectively, in bovine mammary tissue from Chinese Holstein dairy cows [17].
The cytoskeleton plays an important role in maintaining cellular integrity, structure, and function and has been reported to express different cytoskeletal proteins depending on cell type [27]. Therefore, cytoskeletal protein markers such as CK18, CK19, vimentin, and α-SMA were used to establish the primary bovine mammary cells [24,26]. According to previous studies, CK18 and CK19 are characteristic markers for mammary secretory ECs and vimentin and α-SMA are typical markers for mammary FBs and MCs, respectively [28,29]. Consistent with the previous studies, our results showed that CK18 and CK19 were primarily expressed in ECs, and vimentin and α-SMA were mainly expressed in FBs and MCs. However, the differences in the fluorescence and protein levels of cytoskeletons among FBs, MCs, and ECs gradually decreased over the serial passages (Figs. 2 and 3). In fact, primary bovine mammary cells have the finite lifespan along with the instability of passage [15,16,20]. This cellular senescence of primary bovine mammary cells induces an irreversible arrest of cell growth and proliferation, ultimately resulting in cell death along with the occurrence of vacuoles [16]. Therefore, the growth curves of FBs, MCs, and ECs were investigated to evaluate the correlation between cytoskeletal protein and cell numbers and our finding showed that the cell number of these cells gradually decreased over the serial passages (Fig. 4). These results suggest that the cause of the decreased cytoskeletal proteins is directly related to the replicative senescence. Taken together, our data indicate that the cytoskeleton characteristics were not maintained due to a decrease in cell numbers resulting from cellular senescence over serial passages.
Milk synthesis and secretion in ECs are primarily regulated by the coordinated action of reproductive hormones (PRL, P4, and E2) and metabolic hormones (INS and CORT) [29]. PRL is a key hormone that promotes alveolar differentiation and milk component production [30]. Therefore, in the current study, ECs were differentiated using DCM supplemented with various hormones, including PRL, to evaluate the milk component productivity through serial passages. The concentration of hormones in DCM was established based on previous studies reporting the production of milk components in ECs [17,31,32]. ECs differentiated with DCM at passage 1 significantly increased milk protein-related genes and milk fat-related triglycerides compared to those that proliferated with CCM (Fig. 5). However, the synthesis of milk proteins and fats gradually decreased with an increase in vacuoles in ECs over serial passaging. According to a previous study, the occurrence of large cytoplasmic vacuoles and the decrease of milk component productivity were the major feature of involution in ECs of mammary gland [33]. In particular, involution of the mammary gland is known to associated with the impairment of TJs integrity as a process of returning milk-secreting ECs to their non-lactating state [26,33]. Thus, the correlation between milk component productivity and TJs integrity, depending on hormone addition, was further investigated using CCM and DCM. Previous studies have reported that mammary gland involution occurs in two distinct physiological phases in the absence of lactogenic hormone secretion [30,34]. Early apoptosis and the loss of TJs occurred in the first phase, and anoikis was irreversibly induced with a massive loss of mammary ECs after disruption of the basement membrane through the activation of MMPs in the second phase [35]. In contrast, the presence of hormones, such as PRL and CORT, enhanced the TJs formation and milk production by mammary ECs [36]. Indeed, culture media supplemented with various hormones such as PRL, INS, CORT, P4, E2, and EGF induced and maintained milk component production and cellular characteristics in spontaneously immortalized primary yak and buffalo mammary ECs up to passages 50 and 60 [29,37]. In line with these earlier findings, the present study demonstrated that CCM induced involution and the formation of cellular vacuoles, whereas DCM led to differentiation and the development of alveolar morphology. Moreover, ECs cultured with CCM could not synthesize milk components because of the impairment of TJs caused by MMP activation, whereas DCM elevated milk component production by enhancing TJs and decreasing MMPs activity. Taken together, the absence of hormones in CCM resulted in MMP-induced impairment of TJs integrity, and the presence of hormones enhanced milk component productivity and TJs protein expression via a decrease in MMP activity in ECs. Therefore, our data suggest that hormones are essential for maintaining both cellular characteristics and milk component productivity in ECs.
Several prior studies have successfully established mammary gland EC models to investigate their functions and milk synthesis [17,18,38]. Nonetheless, these studies have predominantly focused on assessing the cellular characteristics of ECs during their early passages. Consequently, our research holds significance in elucidating the impact of hormones and the factors contributing to cellular instability as we are aiming to establish and maintain cellular stability across successive passages. Additionally, among the array of hormones, while PRL plays a pivotal role in determining cellular stability in ECs [39], its application is constrained by the prohibitive cost of recombinant PRL [40,41]. Consequently, there is a need for further research into alternative PRL substitutes to facilitate the production of cell-cultured milk components by ensuring the establishment of cellular stability in ECs.
In conclusion, our study demonstrated that primary bovine mammary cells in their early passages have high expression in cytoskeleton (CK18, CK19, vimentin, and α-SMA) and milk components (CSN1S1, CSN2, CSN3, ALA, BLG, and triglycerides)-related markers. These results indicate that primary bovine mammary cells have the cellular stability in the structure, function, bioactivity, and bioproduction at early passages. However, these cellular characteristics and functions gradually declined in subsequent passages. Additionally, we observed that primary bovine mammary ECs exhibited decreased milk component production in the absence of hormones, attributed to the damage induced in TJs by MMP. Conversely, the introduction of various hormones into primary bovine mammary ECs resulted in increased milk component productivity and preserved TJs integrity by inhibiting MMP activity. In summary, the establishment of cellular stability in primary bovine mammary cells depends on the coordinated action of diverse hormones. To facilitate the production of cell-cultured milk in cellular agriculture based on animal cells, the optimization of hormone use and exploration of potential PRL substitutes are imperative.