
INTRODUCTION
Pork feed primarily consists of soybean meal and corn, supplemented with various additives such as vitamins and minerals to control the growth rate of pigs [1,2]. The composition of pig feed can also influence pork quality [3]. Many studies have been conducted to improve both pork production and quality by supplementing pig feed with various additives, including antioxidants [4]. Vitamin C, vitamin E, and selenium (Se) have been used as antioxidants in feed, and previous research has shown that their use can modulate the antioxidant capacity, nutritional quality, and fatty acid composition of pork [1,5].
Se is a commonly used in pork farming due to its regulatory and immune system function [6,7]. It can also improve pork quality and nutritional value as it is an essential components of glutathione peroxidase (GPx) [8,9]. GPx is one of the antioxidant enzymes that can reduce lipid hydroperoxides and free hydrogen peroxide in body tissues [10]. Therefore, Se supplementation can increase GPx activity, potentially improving antioxidant capacity of pork [11].
Se exists in two chemical forms in nature, organic and inorganic [12]. Inorganic Se, mainly in the form of selenite and selenium salts, is commonly used in pork feed due to its easy supply and cost-effectiveness [13]. However, the use of inorganic Se has limitations such as low accumulation rate in the body despite high digestion and absorption rate [14], lower absorption rate compared to organic Se [15], and potential toxic effects at high levels [16].
On the other hand, organic Se, in the form of selenomethionine and selenium-yeast, has a higher accumulation efficiency and antioxidant activity when fed to livestock [17,18]. It can also prevent Se deficiency, which frequently occurs in weaning piglets when fed to sows [19]. In addition, organic Se has been reported to delay the post-oxidative reaction of the muscle, improving the nutritional value, flavor, and shelf life of meat, as well as meat color and water holding capacity (WHC) [20–22]. Despite being expensive, organic Se has been considered for pig feeding [23].
Recently, there has been emphasis on converting feed supplements from inorganic Se to organic Se due to the limitation of Se and the potential benefits of organic Se [24]. However, economic feasibility is an important factor in livestock industry, and the conversion rate must be considered. Several studies are currently underway to replace and/or combine inorganic Se with organic Se, and some have reported improved antioxidant performance and health levels [25]. While we have confirmed the combined effect of inorganic and organic Se on the growth performance of pigs at different levels (data not shown), their effect on antioxidant capacity and quality has not been studied for our market consumers. Therefore, we evaluated the combined effect of inorganic and organic Se on the quality of pork loin during refrigerated storage.
MATERIALS AND METHODS
A total of 105 growing pigs ([Yorkshire × Landrace] × Duroc) with an average body weight of 39.85 ± 0.01kg were divided into 15 pens with 7 pigs in a randomized complete block design. The pigs were kept in climate-controlled facility that had a fully concrete floor measuring 2.4 by 2.9 m2. A feeder and a nipple drinker were provided in each pen to ensure that the pigs had unrestricted access to food and water. The experimental period was 14 weeks during with three types of experimental treatments were implemented. Each of the 5 pens was assigned to one of 3 treatment groups, resulting 5 pens per group. The experimental treatments were as follows: Con (fed basal diet), Se15 (fed 0.15 ppm organic Se along with 0.10 ppm inorganic Se), and Se45 (fed 0.45 ppm organic Se along with 0.10 ppm inorganic Se). Each treatment group was fed with 0.10 ppm of inorganic Se (Genebiotech, Gongju, Korea), while the addition of organic Se (Sel-PlexTM, Alltechm, Nicholasville, KY, USA) was adjusted to induce Se accumulation in pork. The transformation from inorganic to organic Se was accomplished by partially modifying the feeding quantity of inorganic Se. From each group, 5 pigs were randomly selected and their loins (M. longissimus) were obtained. The samples were cut into 3 pieces (330 ± 20 g) and packaged in air permeable bags. They were then stored at 4°C, and the following experiments were conducted on days 0, 7 and 14. On each storage day, WHC, pH, and meat color were analyzed immediately, and the samples were frozen at −70°C until further analyses.
The Se concentration in pork loins was determined using the fluorometric method. To perform the analysis, 0.5 g of the sample was added to a screw cap culture tube containing 5 mL of a mixed solution of HClO4 (perchloric acid 70%) and HNO3 (nitric acid 70%) in 1:4 ratio. The culture tube was digested for 4 h in a digestion block at 210°C, then cooled down in room temperature. After cooling, add 0.5 mL HCl was added to the tube and the tube was heated at 150°C for 30 min. Then, the tube was cooled again, and 15 mL of 0.1M EDTA solution and 2 mL of 0.1% 2,3-diaminonaphthalene solution were added. The tube was voltexed for 5 sec and incubate in a water bath at 60°C for 30 min. Following incubation, a 10-second vortexing of the tube was done after adding 5 mL of cyclohexane. The extracted cyclohexane layer was transferred to a cuvette, and the absorbance was measured using 369 nm excitation and 525 nm emission settings.
The activity of GPx activity was measured through the utilization of Glutathione Peroxidase Assay Kit (353919, Sigma-Aldrich, Burlington, USA). Briefly, minced meat sample (5 g) was homogenized with 25 mL of cold homogenization buffer (50 mM Tris-HCl, pH 7.5, 5 mM EDTA, 1 mM DTT) at 12,000 rpm for 1 min (T25 digital ULTRA-TURRAX®, IKA, Staufen, Germany). The homogenized sample was centrifuged (Continent 512 R, Hanil, Incheon, Korea) at 10,000×g for 15 min, and the supernatant was taken. The Assay Buffer, Co-Substrate Mixture, and NADPH included in the kit were mixed with the supernatant. Then, the reaction was initiated by adding hydroperoxide. Thereafter, the absorbance was measured at 340 nm every min for 10 min to confirm the GPx activity.
Ground sample (3 g) was homogenized with 12 mL of deionized distilled water at 9,600 rpm for 30 s (T25 digital ULTRA-TURRAX®, IKA). The homogenized samples were centrifuged (Continent 512 R, Hanil) at 2,265×g for 10 min, and filtered using filter paper (No. 1, Whatman PLC, Maidstone, UK). For the meat extract, after centrifuging at 2,265×g for 10 min, 10 mL of chloroform was added to the filtrate.
For the 2,2′-azinobis-(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) assay, a solution of 14 mM ABTS and 4.9 mM potassium persulfate was prepared and left in the dark for 16 minutes after vigorous vortexing. The subsequent steps were performed following the protocol described by Choe et al. [26].
For the 2,2-diphenyl-1-picrylhydrazyl (DPPH) assay, 1 mL of the diluted meat extract was mixed with 1 ml of 0.2 mM DPPH in methanol, vortexed, and placed in the dark for 30 min at room temperature. The subsequent steps were performed following the protocol described by Choe et al. [26].
For the 2-thiobarbituric acid reactive substances (TBARS) assay, the meat sample (5 g) was homogenized with 15 mL of deionized distilled water and 50 μL of 7.2% butylated hydroxy toluene solution at 9,600 rpm for 30 s (T25 digital ULTRA-TURRAX®, IKA). Then, the subsequent steps were followed by Rupasinghe et al. [27].
Minced meat sample (5 g) was placed on a filter paper and centrifuged at 252×g for 10 min (Continent 512R, Hanil). The WHC was measured as described by Kwon et al. [28] and pH by Rupasinghe et al. [27], respectively. The meat color of pork loin was measured using a colorimeter (CM-5, Konica Minolta, Osaka, Japan). Prior to measurement, the colorimeter was calibrated with a standard black plate. The meat color was measured at three different locations on the top and the bottom of each sample [22]. The color value was expressed as CIE L*, CIE a*, CIE b* and delta E was calculated as.
The NMR analysis was performed according to Kim et al. [29]. In brief, each minced sample (5 g) was homogenized with 20 mL of 0.6 M perchloric acid at 12,000 rpm for 1 min (T25 digital ULTRA-TURRAX®, IKA). The homogenized samples were centrifuged at 2,265×g for 20 min (Continent 512R, Hanil), and the supernatant was transferred in another test tube and adjusted to 7.0 with sodium hydroxide. Then, the subsequent steps were performed following the method [29].
The data were analyzed using two-way analysis of variance (SAS 9.4, SAS Institute, Cary, NC, USA). The mean values and standard errors of the means were presented as the results. Differences with a significance level of 0.05 were determined by the Student-Newman-Keuls multiple range test. Partial least squares-discriminant analysis was conducted using MetaboAnalyst 5.0 (https://www.metaboanalyst.ca).
RESULTS AND DISCUSSION
Throughout all storage days, the pork loin supplemented with Se45 showed the highest Se contents followed by Se15 and Con (Fig. 1; p = 0.0009). This indicates that the higher organic Se supplementation leads to higher residual Se contents in pork loins, as organic Se sources are highly bioavailable [15,29]. When Se-yeast was supplied as organic Se source, the amount of Se in the loin increased with increasing Se concentration in the feed [30]. Zhan et al. [22] also confirmed that pig muscle Se content increased more than double when fed with organic Se compared to inorganic Se. According to the findings of Zhang et al. [31], intramuscular Se content increased significantly when SeMet was used as a feed source, in comparison to inorganic Se sources such as SeNa or basic feeding treatment groups. Furthermore, organic Se has low toxicity, high transfer efficiency, and the ability to build and maintain Se reserve in muscle [30].
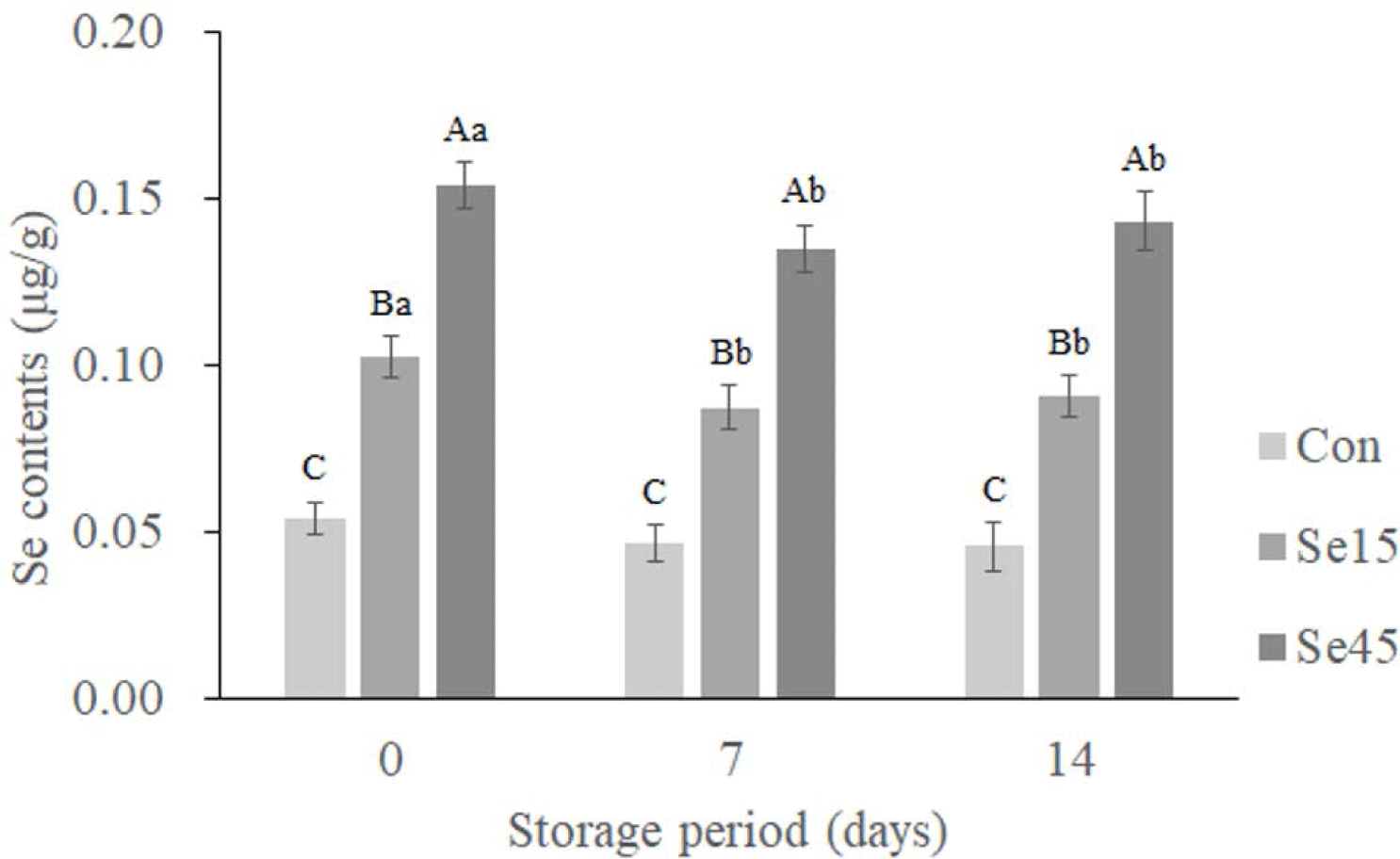
Meanwhile, Se contents were slightly decreased in Se15 and Se45 on day 7 and remained constant thereafter (Fig. 1; p < 0.0001). This reduction in Se content in pork during the refrigerated storage is likely due to microbial activity, temperature, etc. [32]. Despite this decrease, Se15 and Se45 still had higher Se contents than Con, indicating that the effect of Se supplementation can be maintained in pork during storage. We found no further impact from the interaction between the treatment and storage period (p = 0.6826).
The increased Se content in pork can have various impacts, as Se may have prevented oxidative damage from live animals to meat storage [33]. Therefore, high productivity can be promoted for pigs, consumers who lack selenium can be relieved, and several beneficial effects can be provided to consumers. Se supplementation in live animals can improve reproductive physiological characteristics, such as semen volume and semen concentration [34]. Furthermore, Se supplementation in live animals can improve reproductive physiological characteristics, such as semen volume and semen concentration [34]. Furthermore, Se content in milk from sow increases, which has the advantage of solving Se deficiency that can easily occur in piglets [19]. With regards to meat quality, the supplementation of organic Se can enhance meat color stability by protecting myoglobin from oxidation with its antioxidant ability [22]. Calvo et al. [35] confirmed that Se-fed pork has high lipid stability during storage. In addition, consumption of Se-enriched pork may result in a reduction in toxic factors, as Se in pork has the ability to bind with heavy metals (such as cadmium, mercury, zinc, etc.) and facilitate their excretion from the body [36,37]. Moreover, Se content in pork exhibits antioxidant effects by interacting with various antioxidant enzymes in the body, which can prevent DNA damage by averting several harmful effects of free radicals [38]. Therefore, when higher organic Se is fed to pigs, pork with the higher Se content can be served to consumers, providing additional health benefits at the point of their consumption.
GPx is an antioxidant enzyme that contains Se [8,9] and can be increased by Se supplementation in pigs [39,40]. As a result of confirming GPx activity in this study, organic Se supplementation had a significant effect on GPx activity (Fig. 2; p = 0.0179), but the effect of interaction between organic Se supplementation and storage period was not confirmed (p = 0.7874). Previous research has indicated that selenium can be absorbed through the digestive system and subsequently accumulated in various organs [6]. The accumulated Se undergoes various metabolic processes and plays a key role in the synthesis of GPx. As GPx contains Se in its active center, increased uptake and accumulation of Se in the body can promote its activity [41].
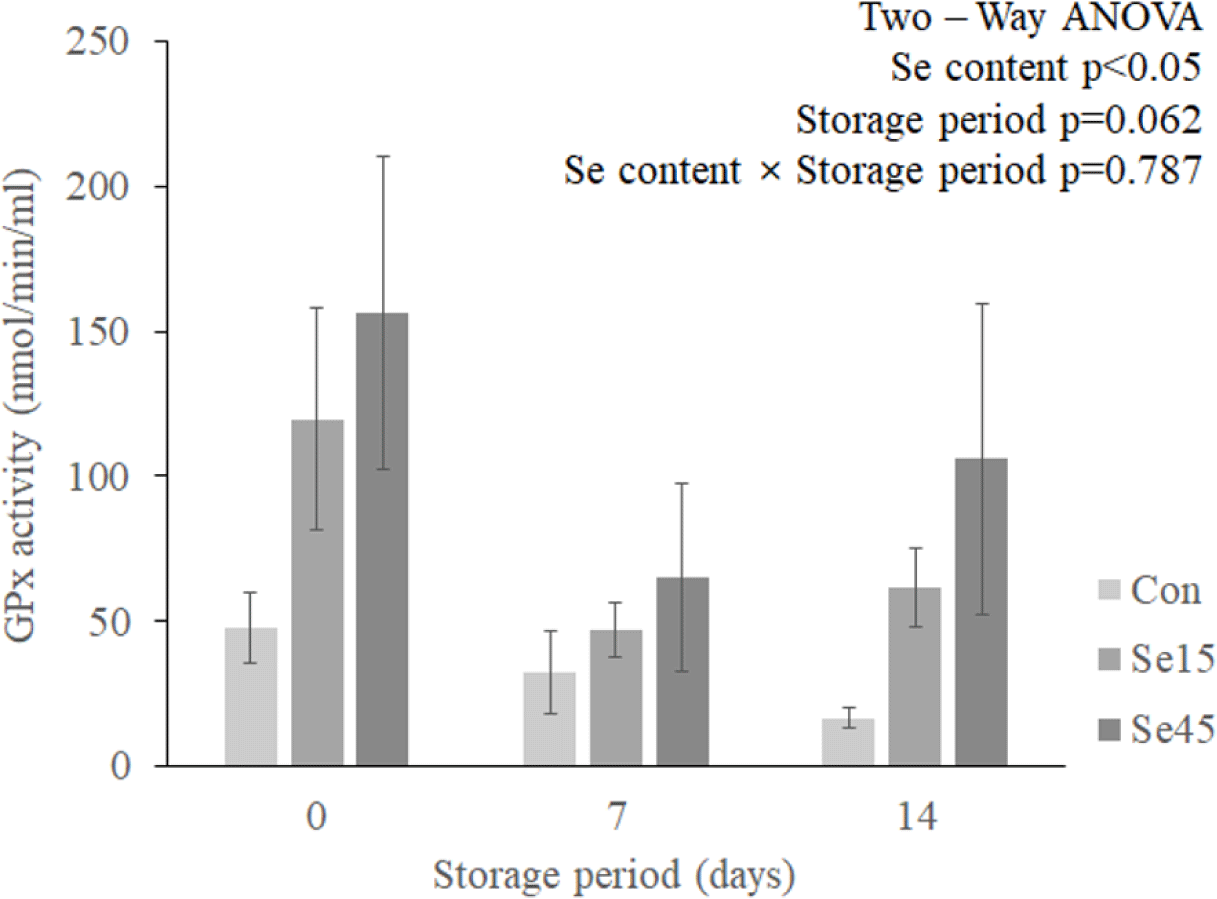
The increased activity of antioxidant enzymes may improve the storage stability of meat. Although the Se content in muscle decreased as the storage days increased in the experimental groups fed Se, Se45 had the highest Se content on all storage days. The increased activity of antioxidant enzymes can increase the antioxidant capacity of meat, which can have a positive effect on improving meat quality such as storage stability.
To investigate antioxidant capacity of Se-supplemented pork, ABTS/DPPH scavenging activities were conducted (Table 1). Organic Se supplementation did not significantly change the ABTS scavenging activity, the DPPH scavenging activity showed a similar trend in each treatment, possibly due to their strong correlation (r = 0.906). These unexpected results could be attributed to the fact that the change in GPx activity was not sufficient to affect the antioxidant activity of meat (Figs. 1 and 2). Although GPx plays a role in reducing lipid peroxide to alcohol and free hydrogen peroxide to water [10], ABTS/DPPH scavenging activities confirm the antioxidant effect through scavenging of free radicals, not hydrogen peroxide, and may not directly related to the high activity of GPx.
During 14 days of storage period, the tendencies in DPPH and ABTS scavenging activities were different (Table 1). ABTS scavenging activity was gradually increased, possibly due to the increased functional peptides from protein degradation during post-mortem (p < 0.05) [42]. However, in the case of DPPH assay, its activity was significantly decreased on day 7 and increased thereafter. The different results in ABTS and DPPH scavenging activities may be attributed to different mechanisms and subjects of both analytical methods. The ABTS assay is for both hydrophilic and lipophilic antioxidants, whereas DPPH assay is more applicable to hydrophobic system. It seems that post-mortem changes in pork induced stronger impact on ABTS and DPPH scavenging activities than that from organic Se supplementation.
Lipid oxidation is a major concern in pork quality, as it can negatively affect acceptability of the meat. The oxidation of lipids can occur due to the inadequate scavenging capacity of antioxidants against the release of free radicals [43]. The extent of lipid oxidation during storage was assessed by conducting TBARS analysis as shown in Table 1. In the present study, organic Se supplementation did not exhibit a significant impact on lipid oxidation compared to the control group. This was unexpected as meat GPx activity can counteract free radicals, thereby influencing lipid oxidation [44]. Several factors may have contributed to this finding. Firstly, slow lipid oxidation rate by low-fat content in pork loin may have made it difficult to observe the differences from the enhanced GPx activity in the Se-supplemented groups (Fig. 2), as fat content is one of the main factors affecting lipid oxidation [43]. Additionally, the progress of lipid oxidation may have been delayed as the samples were stored at low temperatures. Consequently, we found that the lipid oxidation barely occurred in all groups after 14 days of storage, regardless of different Se feedings (Table 1). On day 7, a slight but significant decrease in TBARS value was found only in the Se-supplemented groups. Secondly, the increase in GPx may not have been enough to inhibit further lipid oxidation in pork loin. Hoac et al. [45] reported a certain decrease in lipid oxidation by GPx activity when 4 U/g GPx was added to chickens and ducks.
Taken the results from antioxidant properties together, although Se supplementation improved the activity of GPx, these changes did not affect the antioxidant activity and the lipid stability of pork loin during storage.
During the storage period, no significant difference was observed in WHC and pH between the control and groups supplemented with organic Se (Table 2; p = 0.5897 and p = 0.2557, respectively). However, the changes in these properties varied depending on the levels of organic Se supplementation. During 14 days of storage, the WHC changed by 13.59%, 18.79%, and 18.89% in the control, Se15, and Se45 groups, respectively. It can be attributed to the decrease in water content over time (data not shown), as its decrease may limit free water release [46]. Similarly, the pH decreased at different rates in each group, with the control group having a decrease of 0.39, while Se15 and Se45 had reduction of 0.26 and 0.24, respectively. Even though several studies have reported that organic Se supplementation can increase WHC and reduce the decrease in pH in pork after slaughter [33,47], in this study, organic Se supplementation (15 or 45 ppm) with 10 ppm inorganic Se did not affect WHC and pH in pork during 14 days of storage.
In regards to meat color, there was no significant difference in the CIE L*-, CIE a*-, and CIE b*-values among different organic Se supplementation, except for CIE a*-value on day 7 (Table 3). While previous studies have reported that organic Se supplementation at 0.3 ppm can increase CIE a* and CIE b*-values [35], this study did not observe any changes in meat color due to the lack of pH change in pork. The pH plays an important role in the mechanism by which oxymyoglobin is oxidized to metmyoglobin. In the case of Se-yeast, a type of organic Se fed in this experiment, it was absorbed through the methionine transporter and incorporated into the protein constituting the body, suggesting that it may not have affected meat quality, including its color. Nevertheless, previous research has indicated that consumption of organic Se may enhance muscle antioxidant capacity, protecting myoglobin from oxidation and thereby improving color stability [22]. Conversely, inorganic Se has been reported to induce lighter color than pigs fed with organic Se, mainly due to water droplet loss that occurred when fed with inorganic Se [21].
During storage, different atmospheres can cause variation in the meat color of pork can [46]. The total color difference (ΔE) was calculated to confirm the changes in color (Table 3). Overall, no distinct color changes were observed in this study, indicating that the organic Se supplementation did not affect meat color in pork loin. The CIE L*-value tended to decrease, possibly due to an increase in WHC (Table 2), regardless of the type of organic Se supplementation. The CIE a*-value in each group was also affected by post-mortem changes. Its increases on day 7 is possibly due to the oxygenation of myoglobin and the value decreased due to oxidation to metmyoglobin [48]. No previous study has investigated the effect of mixed feeding of organic and inorganic Se on the meat color of pork. Based on the results of this study, the organic Se supplementation treatment did not affect meat color.
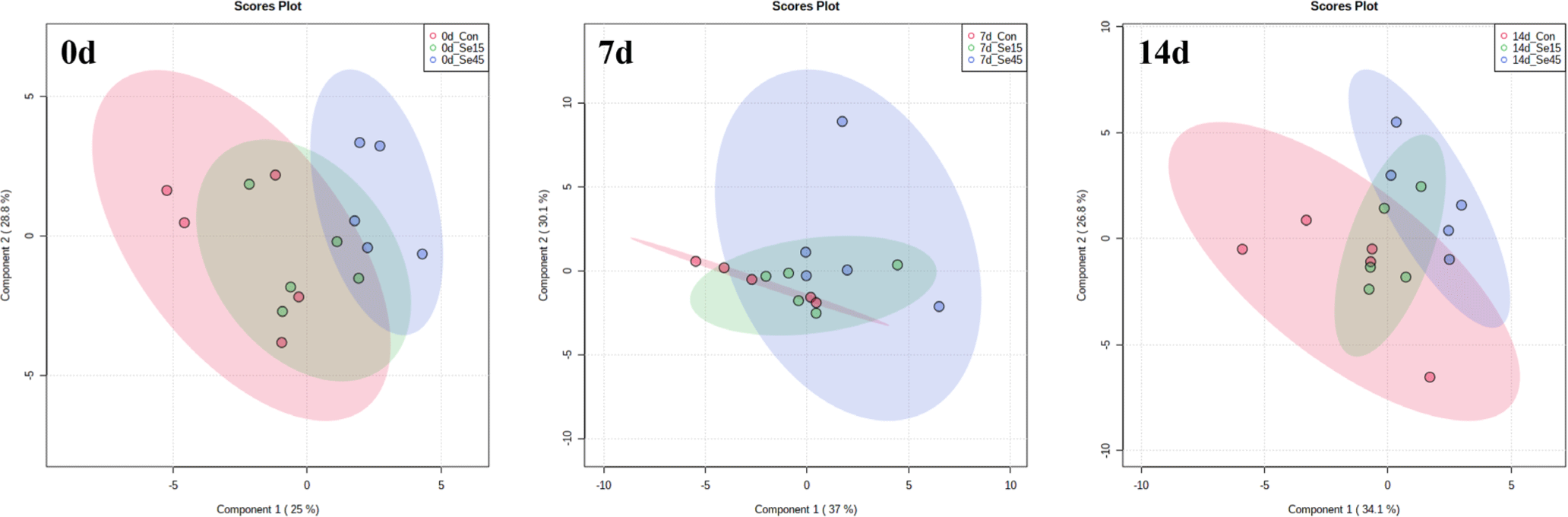
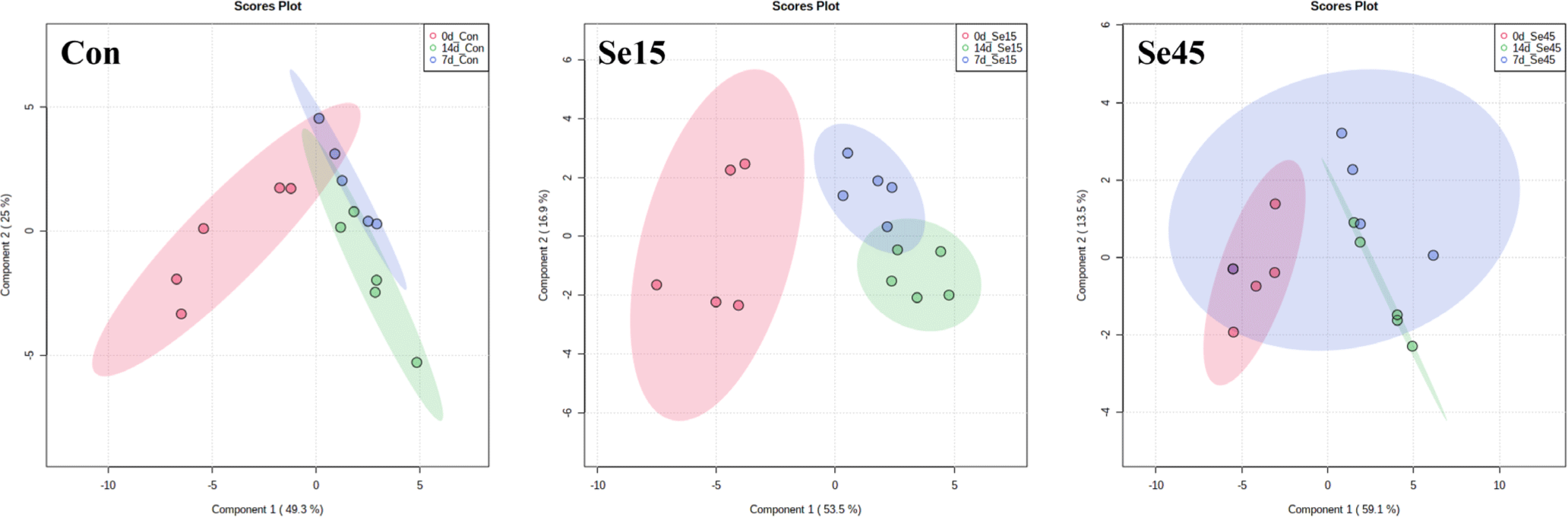
We performed NMR-based metabolic analysis to investigate the effects of different Se supplementation on the metabolic profiles of pork loin during 14 days of storage. Table 4 presents a total of 31 metabolites that were identified across all groups, including 15 free amino acids, 4 nucleotide-related products, and 3 organic acids. To assess the metabolomic differences among treatment groups and storage periods, multivariate analysis was performed, as shown in Figs. 3 and 4, respectively. The metabolic profiles of Con, Se15, and Se45 were not distinctly different from each other on each storage day, as indicated in Fig. 3. This suggests that the accumulated Se content in Se15 and Se45 did not have an impact on the metabolic differences during the storage period. No significant changes in metabolites, except for a few such as tyrosine, inosine, and betaine on day 0 and glutamate on day 14, were observed with different Se supplementation. Furthermore, lactate content was not significantly different between Con and both Se-supplemented groups (Table 4), but its content increased during storage, leading to a pH decrease (Table 2). Although slight changes in the metabolites in each group were observed during storage period, in overall, these changes were not distinct (Fig. 4). Each group exhibited different changes in the levels of amino acids (alanine, asparagine, creatine, glutamate, glutamine, glycine, isoleucine, leucine, methionine, phenylalanine, threonine, tyrosine, and valine) and nucleotide-related compounds (hypoxanthine and inosine), as shown in Table 4. These changes can be attributed to the degradation of proteins and nucleic acids during storage, leading to an increase in the content of degradation products [49]. Additionally, lactate, which was previously mentioned, the other metabolites (acetate, carnosine, ethanol, glucose, N,N-dimethylglycine, niacinamide, and O-acetylcarnitine) also showed significant changes during 14 days of storage, but not due to Se supplementation. These results suggest that the Se feeding conditions used in this experiment were not sufficient to induce metabolomic changes in pork loin.
CONCLUSION
This study found that different levels of organic Se (0.15 and 0.45 ppm) combined with inorganic Se did not significantly affect pork quality during 14 days of storage, despite an increase in tissue Se content and GPx activity. Therefore, high Se content in the organic Se-fed group may have a positive effect on Se accumulation in pig muscle, but organic Se supplementation up to 45 ppm does not affect pork quality during storage periods of up to 14 days. In the results of supplementation with Se, the same phenomenon as the control group was confirmed on all days of storage. Therefore, through this study, it was confirmed that Se, a trace mineral used for pig breeding management, does not adversely affect pork quality.