
INTRODUCTION
Sperm is the male germ cells which deliver the haploid paternal genome to the eggs to generate offspring. In mammals, sperm are ejaculated into female reproductive tract and migrate toward the fertilizing site using their cilia-like sub-cellular organelle, flagella [1,2]. Although this hair-like organelle could be simply considered to generate mechanical force for sperm navigation in female tract, complex cellular signaling pathways determine flagellar movement and sperm swimming pattern [3,4]. In female reproductive tract, sperm sense the extracellular environment and initiate various signaling pathways [5]. Eventually, the activated signaling pathways in responding to the environment changes physiological characteristics of sperm to fertilize eggs, which is the process called capacitation [6,7]. Interestingly, the capacitated sperm change their motility pattern, characterized by asymmetric flagellar beating with increased amplitude, called hyperactivated motility [8]. By developing this unique swimming strategy, mammalian sperm successfully reach the fertilizing site and penetrate the eggs’ barrier, zona pellucida, followed by fertilization.
Hyperactivated motility is achieved by influx of extracellular Ca2+ into the sperm via sperm-specific cation channel, CatSper [9,10]. CatSper is one of the most complex ion channels composed of at least 14 subunits. Previous studies using mouse models identified four six-transmembrane (TM) subunits, Catsper1–4 [9,11,12], to form heterotetrameric pore; single TM subunits, CatSperβ, γ, δ, and ε [13–16] to form the huge extracellular canopy structure; and cytoplasmic subunits, CatSperζ and EFCAB9 [16,17]. In addition, recent studies to resolve atomic structure of the CatSper channel further report CatSperθ (TMEM249), CatSperη (TMEM262), and Slco6C1 as the CatSper components [18, 19]. The CatSper complexes arrange quad-linearly to form unique Ca2+ signaling domain on the sperm principal piece [16,17,19,20]. With the unique domain structure, the CatSper channel serves as the major Ca2+ entry site in sperm to develop hyperactivated motility in mammalian sperm. Thus, the CatSper deficiency or its altered function cause defective sperm hyperactivation and impaired male fertility in human and mouse [5]. However, the physiological significance of the CatSper channel in sperm Ca2+ signaling and hyperactivation is not well understood in other mammals including domestic animals.
CatSper subunits are even conserved in unicellular flagellates [21]. Although the subunits are lineage-specifically conserved in certain animals [16,17,22], their functional regulation and activation mechanisms seem to be variable within species. In mouse, CatSper channel is mainly activated by the intracellular alkalinization [23]. By contrast, although intracellular alkalinization can contribute to activating CatSper channel [24], an additional ligand, progesterone, is further required to fully activate the CatSper in human [25,26]. The difference between human and mouse sperm suggests the species-specific characteristics of the CatSper channel in mammalian sperm. Yet, our knowledge of the mammalian CatSper channel is only limited to human and mouse sperm, which requires more insight into the CatSper channel in other mammals. In that context, domestic animals could be good models to expand our knowledge for CatSper channel in mammals due to their commercially available semen samples.
In this review, I briefly introduce sperm hyperactivation by CatSper channel and walk through what was previously known for the CatSper channel in domestic animals with comparative aspect.
In female reproductive tract, ejaculated mammalian sperm gradually acquire the fertilizing ability via capacitation [6,7]. Interestingly, ejaculated sperm retrieved from the fertilizing site, ampulla, have vigorous flagellar beating with increased amplitude distinguished from epididymal sperm [8]. This motility pattern is also triggered by incubation with follicular fluid in vitro [27]. The changed motility pattern of mammalian sperm by female factors are now termed hyperactivated motility. After the first observation of the hyperactivated motility in hamster sperm, this motility pattern has been also observed from sperm from various mammals [28]. Hyperactivated motility provides increased mechanical force to sperm. Thus, by developing this unique motility pattern, sperm can successfully pass the lumen filled with mucus with escaping the oviductal reservoir [29,30]. In addition, sperm require hyperactivated motility to penetrate zona pellucida followed by fertilizing eggs after their arrival to the fertilizing site [31]. Therefore, hyperactivated motility is indispensable for successful migration and fertilization of the mammalian sperm in female tract.
Although follicular fluids can induce sperm hyperactivation in vitro [27], downstream signaling pathway to develop hyperactivated motility had not been well understood. Later, individual factors, such as bicarbonate, cAMP, and Ca2+, have been examined to test whether they are the direct physiological triggers to induce sperm hyperactivation. Finally, it was turned out that Ca2+ is the fundamental trigger for sperm hyperactivation in mammals. Sperm fail to develop hyperactivated motility in absence of the extracellular Ca2+ during in vitro capacitation [32,33]. In opposite, treating Ca2+ ionophore (A23187) transiently induces hyperactivated motility in boar and golden hamster sperm [33]. The Ca2+ requirement to develop hyperactivated motility was further supported by demembranated sperm of which flagella form huge arch by the Ca2+ exposure [34]. Furthermore, sperm to develop hyperactivated sperm have relatively higher intracellular Ca2+ levels compared to the non-hyperactivated sperm [10]. All these results clearly demonstrate that sperm develop hyperactivated motility by the increased intracellular Ca2+ level, which are sourced from the extracellular environment. Although Ca2+ from intracellular storage sites, such as mitochondria, has been suggested to develop hyperactivated motility [35], its physiological triggers in female tract, which secret Ca2+ into the sperm cytoplasm, is unidentified yet.
The requirement of the extracellular Ca2+ for sperm hyperactivation indicates directly that sperm would carry Ca2+ channel to introduce extracellular Ca2+ into their cytoplasm. Various voltage-gated Ca2+ channels and transient receptor potential channels had been suggested for the Ca2+ channel for sperm hyperactivation [36-38]. However, those channels are not considered for the major Ca2+ entry site because of their marginal significance in sperm hyperactivation [39]. In 2001, the sperm-specific Ca2+ channel, CatSper, has been identified which is now known for the major Ca2+ channel for sperm hyperactivation [9]. The first identified CatSper subunit, CatSper1, has six TM, like the pore subunits of the voltage-dependent K+ channel. Previous patch-clamping analysis recorded Ca2+ current which is not present in CatSper1-null sperm [23]. These studies demonstrate that CatSper is a Ca2+ channel to introduce the cation into mammalian sperm.
After the first identification of CatSper1, other three six TM CatSper subunits (CatSper2, 3, and 4) were further identified, which form pore of the channel [11,40]. Accumulated studies for the last 20 years reported that CatSper channel is one of the most complex ion channel composed of over 14 subunits [41]. Contrary to the other Ca2+-permeable channels, genetic alterations of the CatSper subunits result in severe defects in male reproduction (Table 1). In mouse, genetic depletion of the CatSper TM subunits results in 100% male infertility in vivo and in vitro [9,12,15,42–44]. Although their absences do not impair spermatogenesis without morphologically abnormal development of the sperm, sperm fail to develop hyperactivated motility. Thus, mouse sperm with impaired CatSper barely pass the utero-tubal junction and arrive at fertilizing site in female tract [16,20,45] and fail to penetrate zona pellucida of eggs [9]. Together with the mouse studies, various genetic mutations in genes encoding CatSper subunits have been identified from infertile male patients (Table 1). Currently, mutations in CATSPER1 [46], CATSPER2 [47–50] and CATSPERE [51] have been identified and the sperm carrying genetic mutation show motility defects. In addition, a missense mutation of CATSPERT was identified from an infertile male with globozoospermia whose sperm motility defect was not confirmed [52]. Thus, currently, CatSper is known for the major Ca2+ channel critical for sperm hyperactivation, which highlights physiological significances of the CatSper channel in male reproduction.
The CatSper is one of the most complex ion channels and previous studies have demonstrated that unique molecular features of the Ca2+ channel is indispensable for sperm hyperactivation in mammals [41].
Notably, CatSper subunits are extremely interdependent in mouse sperm. Although it is still not well understood how the individual subunits are assembled to form functional CatSper channel in developing germ cells, genetic depletion of the CatSper TM subunits results in the absence of the entire CatSper subunits in mature sperm [15–17]. For example, CatSper1- and CatSperd-null mouse sperm do not carry any CatSper subunits although the other CatSper subunits are still expressed in testis [16,17,53]. Considering the partial interaction within the CatSper subunits in absence of the pore subunits [15,53], the CatSper subunits could be only present in the mature sperm tail when they are fully assembled as the functional channel. Interestingly, however, this all-in or all-out of the CatSper subunits in mouse sperm are not observed in human sperm [47]. A previous study detected CatSper subunits in sperm from infertile males who have CATSPER2 mutation [47], which demonstrates the discrepancy for the interdependency within the CatSper subunits in human and mouse sperm.
Interestingly, individual CatSper complexes are linked to each other and arranged on the tapering sperm tail to form unique domain structure [19,20]. CatSper channel localizes specifically at the principal piece of the sperm tail with the quadrilinear arrangement resolved by super-resolution microscopy [20]. Although how the linearly arranged CatSper channels coordinate the spatiotemporal Ca2+ influx into the sperm, the integrity of the domain structure would be crucial for sperm hyperactivation [20,45]. All hyperactivated sperm maintain the domain structure after inducing capacitation in vitro [20] and sperm to arrive at fertilizing site carries intact CatSper arrangement in female tract [45]. These observations strongly support that not only the CatSper channel itself, but its continuous arrangement is required for the successful Ca2+ influx into sperm to develop hyperactivated motility. One possibility is that linearly arranged CatSper would enrich local Ca2+ concentration for the successful sperm hyperactivation like Ca2+ domains in neuronal and immune pre-synapses [54], which requires further studies. This quadrilateral CatSper arrangement is also observed from human sperm [16], which indicates the overall CatSper nanodomain structure could be preserved in mammalian sperm. Recent cryo-electron microscopy (cryo-EM) studies resolved that mouse CatSper channel carries anionic transporter, Slco6C1 [18], which forms the wing-like structure at the zig-zag CatSper domain in mouse [19]. Interestingly, the wing-like structure is not clearly resolved in human sperm [19]. Although this could be simply due to the resolution issue but still there could be differences on the CatSper domain structures between human and mouse sperm, which remains to be further examined.
As mentioned, a few molecular features of the CatSper channel are different between human and mouse sperm despite their common significance to developing hyperactivated motility. Especially, human and mouse sperm requires different activation factors for CatSper channel [23,25,26].
Initially, intracellular alkalinization is known for the key trigger to activate CatSper channel in mouse sperm [23]. Kirichok et al., clearly demonstrated that inward CatSper current is significantly increased by increasing pH in intrapipette solution from patch clamping recording [23]. The CatSper activation by the intracellular alkalinization in mouse sperm is further supported by the other mouse models lacking pH regulators in sperm [5]. The absence of Na+/H+ exchangers, such as sNHE, NHA1, and NHA2, impairs sperm motility with severe fertility defects [55,56]. A previous study demonstrated that the non-TM CatSper subunits, EFCAB9 and CatSperζ, are direct binding partner, and their binary complex serves as pH-dependent Ca2+ sensor to modulate CatSper channel activity [17]. Considering ancient mammals, such as monotremes and marsupials, also carry the EFCAB9 and CatSperζ orthologues (Fig. 1) [57], the pH-dependent CatSper activation and underlying molecular mechanisms to modulate CatSper activity could be conserved in mammals. Protein kinase A signaling by the cyclic AMP (cAMP) was also expected to be another factor to activate CatSper channel by observing increased intracellular Ca2+ level after treating cell-permeable cAMP in mouse sperm [9]. A recent study, however, demonstrated inconsistent effects of the cAMP or cAMP analogues to CatSper activation [58]. Thus, whether the cAMP can activate CatSper channel remains to be clarified in mouse sperm.

As similar to mouse sperm, previous patch clamping studies demonstrated that the CatSper channel can be also activated by intracellular alkalinization in human sperm [24]. Instead of the sNHE in mouse sperm, human sperm carries voltage-dependent proton channel, Hv1, which pumps out the proton from the sperm cells [24,59]. Thus, the proton channel can maintain intracellular alkaline conditions in human sperm to potentiate the CatSper current. However, CatSper channel in human sperm could be further potentiated by steroid ligands, such as progesterone and prostaglandin E1, which is secreted from cumulus cells in female tract [25,26]. Interestingly, different from human sperm, progesterone does not elicit CatSper channel in mouse sperm [9]. In human, α/β hydrolase domain-containing protein 2 (ABHD2) has been suggested as a non-genomic progesterone receptor to activate CatSper channel [60]. ABHD2, a lipid hydrolase, governs unconventional endocannabinoid signaling in a progesterone-dependent manner by depleting 2-arachidonylglycerol, which prevents CatSper activation in human sperm [60]. This progesterone-mediated evoke of the CatSper current is also observed from sperm from macaque monkey [61], suggesting lineage-specific activation mechanisms for CatSper channel in mammal.
Intensive studies in human and mouse sperm have expanded our knowledge of CatSper channel and hyperactivated motility. However, how human and mouse CatSper have different characteristics, especially activation mechanisms, are not well understood. In addition, it is also unclear whether the differences are simply due to the species-specificity of the CatSper or atypical characteristics in either human or mouse sperm. It should be further clarified by expanding our understanding of the CatSper in other species. Thus, domestic animals could be good candidates to explore non-human and non-mouse CatSper with benefits of the easy access of the semen samples. Here, I go over what have been known for the CatSper in sperm from domestic mammals.
After the first observation in Yanagimachi’s study, hyperactivated motility was also reported from bull sperm [62]. In bull, sperm hyperactivation is physiologically significant for not only the successful migration toward fertilizing site, but the dissociation from the epithelial cells at oviduct [63,64]. As like the human and mouse sperm, bull sperm can develop hyperactivated motility by increased intracellular Ca2+ level [34,65,66].
CatSper subunits in bull sperm were identified and characterized first in 2017 [67]. In this study, CatSper1-4 were identified by genomic sequence comparison and CatSper localization at principal piece was confirmed by immunostaining in bull sperm like mouse and human sperm. In addition, the authors demonstrated that blocking CatSper function by treating antibody or Mibefradil, a CatSper inhibitor, significantly reduces sperm hyperactivation and rheotactic response, which is CatSper-dependent in mouse sperm [68]. This study demonstrates the physiological significance of the CatSper in bull sperm hyperactivation.
Interestingly, Johnson et al., also revealed that caffein, which inhibits phosphodiesterase to increase cAMP level, can trigger sperm hyperactivation [67]. This result suggests cAMP signaling pathways could be involved in CatSper activation followed by sperm hyperactivation. One note is that in vitro capacitating medium for domestic animals usually carries ~mM levels of caffein. Thus, to test pharmacological treatment with physiological concentration are still required to confirm significance of the caffein in bull sperm hyperactivation like studies in mouse sperm [58]. Progesterone has been also suggested to activate CatSper channel in bull sperm [69]. Progesterone significantly enhances sperm dissociation from the oviductal epithelium as well as sperm hyperactivation. This was further confirmed reciprocally by treating antagonist for progesterone receptor. Considering its effectiveness at ~nm ranges, like in human CatSper activation, progesterone could be a factor to activate CatSper channel in bull sperm. Yet, its significance in CatSper activation is further examined directly using patch clamping and/or functional imaging assay.
Hyperactivated motility in boar sperm was first reported in 1984 [70]. Influx of extracellular Ca2+ triggers boar sperm hyperactivation, which releases sperm from the oviductal epithelium like bull sperm [71,72].
Porcine CatSper subunits, CatSper1–4, were molecularly cloned in 2011. CatSper1-4 genes highly and/or exclusively express in testis and testicular expression levels increase by sexual maturity [73]. The tissue expression patterns of the CatSper subunits suggest that procedures for CatSper biogenesis in developing germ cells could be conserved in boar. Although the detailed intracellular localization is still unclear, CatSper channel also expresses in ejaculated boar sperm and pharmacological treatment revealed that CatSper inhibition using NNC 55-0396 severely damages boar sperm motility during capacitation [74].
Like bull sperm, progesterone can develop hyperactivated motility in boar sperm, which suggests that progesterone could be also an activator for CatSper channel [75]. Progesterone releases boar sperm bound to glycan-coated beads, which mimic the oviductal epithelium. In addition, to treat a CatSper inhibitor prevents its release from the glycan-coated beads regardless of the progesterone treatment. Furthermore, more directly, Ca2+ imaging assay demonstrated that the intracellular Ca2+ level increases by progesterone, which is blocked by the CatSper inhibitor. All these results elucidate that progesterone is also required for Ca2+ influx via CatSper channel to develop hyperactivated motility in boar sperm. Physiological significance of the cAMP in sperm hyperactivation was also highlighted in boar, but its contribution in CatSper activation requires further studies [76,77].
A previous study characterized CatSper channel in horse sperm [78]. Increasing intracellular pH by NH4Cl enhances curvilinear swimming velocity and amplitude of lateral head displacement in horse sperm, which indicates increased sperm hyperactivation. Interestingly, increasing intracellular pH elevates intracellular Ca2+ level in horse sperm, which is blocked by a CatSper inhibitor, mibefradil. These results suggest that CatSper channel is activated by intracellular alkalinization in horse sperm like the channel in mouse sperm [23]. One note is that progesterone does not affect hyperactivated motility in horse sperm. In addition, another steroid hormone, prostaglandin E1, which potentiates human CatSper current, affects horse sperm hyperactivation neither [78]. All these results indicate that mechanisms for CatSper activation in horse sperm could be distinguished from those in bull and boar sperm.
Although CatSper channel has been intensively understood in mammalian sperm, previous studies reported the homologues for CatSper subunits in early eukaryotes. Comparative genomic studies identified orthologues for the CatSper proteins from early metazoans such as sea squirt [79] and sea anemone [22]. Surprisingly, the sequence homologues were discovered from even unicellular organisms, bikonta and unikonta [21]. In addition, functional study demonstrated that CatSper can introduce Ca2+ into sperm from marine invertebrates, sea urchin [80] and an ascidian - Ciona intestinalis [81], to regulate their motility pattern. These results suggest that CatSper channel could evolve from ancient eukaryotes to modulate flagellar movement by entering extracellular Ca2+.
Interestingly, CatSper subunits seem to undergo distinct evolution in eukaryotes [21]. Comparative genomic studies further revealed that orthologues for all CatSper subunits are not detected from certain lineages, like insects, worms, and mollusks [16,17,22]. By contrast, CatSper components are preserved and functionally conserved as a key Ca2+ channel in earlier metazoans, sea urchin and C. intestinalis [80,81]. This discrepancy highlights lineage-specific evolution of the CatSper channel by all-in or all-out manner. How the CatSper channel is specifically preserved in certain lineages remains to be further understood.
Lineage-specific preservation of the CatSper channel suggests that CatSper components would be evolved rapidly under the strong selective pressure [57]. Indeed, certain CatSper components are newly appeared in specific linage and their orthologues are highly sequence-variable in eukaryotes [16,17,57]. This variation could result in species-specific mechanisms for CatSper activation in eukaryotes.
Despite its conserved role to introduce Ca2+ to modulate flagellar movement, specific chemoattractants, resact and sperm-activating and -attracting factor (SAAF), are required to activate CatSper channel in marine invertebrates [80,81]. By contrast, activation of mammalian CatSper mainly relies on intracellular alkalinization and/or steroid hormone [23,25,26]. This discrepancy within mammals and marine invertebrates could be due to different molecular organization of the CatSper channel. A previous studies identified mammalian-specific CatSper components, CatSperζ [16]. CatSperζ forms a binary complex with another non-TM CatSper subunit, EFCAB9, and the machinery serves as pH-dependent Ca2+ sensor to regulate CatSper activity in mouse sperm [17]. In addition, the interaction between EFCAB9 and CatSperζ are conserved even in ancient mammals [57]. Thus, the newly evolved CatSperζ might determine mammalian-specific mechanisms for CatSper activation in responding to pH-dependent Ca2+ sensitivity.
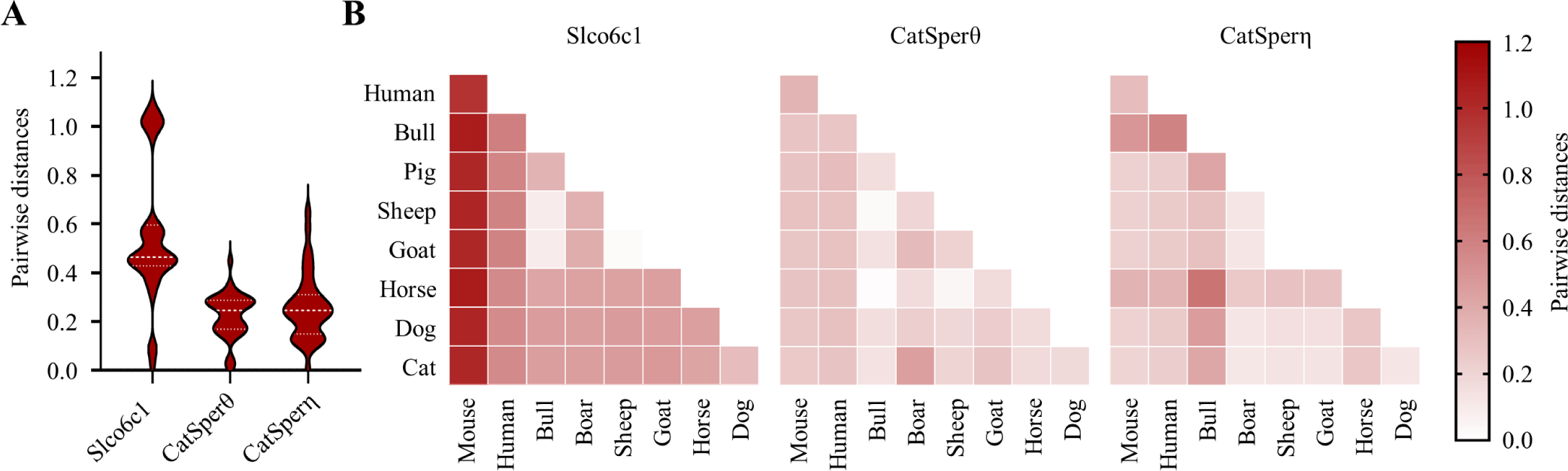
Of note, activation mechanisms of the CatSper channel are even different within mammals. Especially, CatSper activation by progesterone and prostaglandin E1 is critical in human sperm, but not in mouse sperm [41]. Although what causes the difference between two species is still unclear, recent cryo-EM studies to resolve CatSper structure would provide a clue for the question [18,19]. Those studies demonstrated that an organic anion transporter, Slco6c1, is a CatSper component to form wing structure in mouse CatSper complex. Although the ligands and/or substrates for Slco6c1 are not defined yet, Lin et al. [18] speculate that Slco6c1 might mediate steroid hormone transportation. Interestingly, the sequence homologue of mouse Slco6c1 is SLCO6A1 in human, which is also exclusively expressed in testis. Interestingly, Slco6c1 orthologues are only annotated in a few rodents, like mouse and rat, but SLCO6A1 is broadly annotated, indicating the rodent orthologues - Slco6c1 - could have distinctive sequences. Indeed, mouse Slco6c1 is the most sequence variable among the examined sequence homologues in other mammals (Fig. 2). One note is that progesterone triggers Ca2+ influx in sperm from bull and boar, which have orthologues for SLCO6A1. Thus, presumably, the transporters in CatSper complex might determine steroid-mediated channel activation in a species-specific manner, which await to be studied in future.
CONCLUSION
Studies for last 20 years have expanded our knowledge in CatSper-mediated Ca2+ signaling and its physiological significance in male reproduction. Recent structural studies resolved the channel structure in atomic level, which opens the new era for the CatSper channel in translational approaches – developing male contraceptive systems and its application to assistant reproductive technology. However, our knowledge of the CatSper channel has mainly been obtained from mouse models. Despite its great contribution, there are some knowledge gaps between mouse and human CatSper, such as different activation mechanisms. This could be clearly answered by obtaining more information for CatSper channel in other mammals because of its possible species-specificity. Domestic animals could be good models to study CatSper channel because of commercially available semen samples and relatively well-established genome information. Thus, knowledge of the CatSper channel in domestic animals will provide more insights into the CatSper channel with answering remaining questions in future by reducing the current knowledge gaps between human and mouse.