
INTRODUCTION
Ruminants have evolved a complex digestive system to extract nutrients from fibrous plant materials. The diverse community of microbes, including bacteria, protozoa, fungi, and archaea, which inhabit the rumen of ruminants synergistically ferment ingested plant materials, breaking them down into simpler compounds, such as volatile fatty acids (VFAs) [1]. Recent advancements in 16S rRNA gene-based technologies have revolutionized our comprehension of rumen microbial ecology, facilitating the identification and quantification of the diversity and abundance of rumen microbes [2]. To date, most studies have primarily focused on exploring the changes in rumen microbiota associated with dietary changes, methane emission mitigation, heat stress, and feed efficiency [3,4].
Rumen cannulation is a widely adopted, well-established surgical technique for directly obtaining rumen contents, enabling researchers to analyze the composition of rumen microbiota and fermentation characteristics, including VFAs, ammonia, and pH levels [5]. While an alternative approach involving stomach tubing and tube insertion into the esophagus exists, rumen cannulation facilitates the collection of both liquid and solid fractions; moreover, the risk of salivary contamination is lesser than that associated with the stomach tubing method [6,7]. Rumen cannulation has significantly advanced our understanding of ruminal digestion and the intricate interactions between rumen microbes and their ruminant hosts. However, it requires surgical preparation and a postoperative care period of at least 3 weeks, which may potentially induce pain and stress in the animal [8]. Consequently, this procedure can have a considerable impact on the composition of rumen microbiota. Wang et al. [9] observed shifts in the methanogen community following rumen cannulation; however, they did not investigate changes in the rumen bacterial community caused by this procedure.
Hanwoo cattle, the Korean native breed of taurine-type beef cattle, account for over 95% of the total beef cattle population in Korea, comprising approximately 3 million animals [10,11]. With the recent increasing trend of conducting studies concerning rumen microbiota in cannulated Hanwoo cattle using 16S rRNA gene amplicon sequencing, investigating the suitability of the rumen cannulation method for assessing rumen microbiota is essential. Nevertheless, knowledge regarding the effects of rumen cannulation surgery on the composition of rumen microbiota in Hanwoo cattle is limited. Therefore, the present study aims to investigate the longitudinal changes in the rumen microbiota composition of Hanwoo steers that have undergone rumen cannulation surgery by comparing the preoperative and postoperative parameters.
MATERIALS AND METHODS
Eight Hanwoo steers (age, 15 months; initial body weight, 341.4 ± 20.4 kg [means ± standard deviation]) were used in this study. They were fed a total mixed ration (Table 1) [8]. Four steers underwent rumen cannulation surgery (cannulation group), while the remaining four were left intact (control group). The cannulation surgery was performed using 4-inch rumen cannulas (Bar Diamond, Parma, ID, USA) according to a procedure described previously [8]. Rumen samples were collected from the eight steers on the preoperative day (day 0) and on postoperative days 1, 4, 7, 10, 14, 17, 21, 24, and 28, resulting in 80 samples (4 animals × 2 groups × 10 timepoints) using the stomach tubing method, as described previously [7]. The pH of the collected rumen samples was immediately measured using a pH meter (SevenEasy pH, Mettler-Toledo, Greifensee, Switzerland).
Metagenomic DNA from the 80 rumen samples were extracted using the repeated bead beating plus column (RBB+C) method [12]. The DNA samples were used to analyze bacterial compositional shifts related to the cannulation effect by 16S rRNA gene sequencing. The raw data from the 16S rRNA gene amplicon sequences have been deposited in the NCBI Sequence Read Archive under BioProject PRJNA796841.
Bacterial diversity and composition were analyzed using 16S marker gene sequencing according to a previously described protocol [13]. The raw paired-end amplicon sequences, prepared using the 341F (5ʹ-CCTACGGGNGGCWGCAG-3ʹ) and 805R (5ʹ-GACTA CHVGGGTATCTAATCC-3ʹ) universal primers [14], were preprocessed using the DADA2 pipeline [15] implemented in Quantitative Insights into Microbial Ecology version 2 (QIIME2) [16]. The quality-score cutoff was Q > 25. The resulting amplicon sequence variants (ASVs) were taxonomically classified using a naïve Bayes taxonomy classifier trained on the latest Silva database (NR 138 version) [17]. Any reads classified as eukaryotes, mitochondria, or chloroplasts were removed from the resulting high-quality ASVs before downstream analysis.
Alpha diversity indices (observed ASVs, Chao1 estimates, Shannon’s diversity, and Simpson’s diversity) were computed to analyze richness, evenness, and phylogenetic diversity differences between the control and cannulated groups. These analyses were conducted using rarefied tables with randomly chosen 40 997 ASVs for each sample. Good’s coverage exceeded 99.4% across all samples. The overall distribution of bacteriota was examined using principal coordinates analysis (PCoA) based on unweighted and weighted UniFrac distances. These results were visualized using the emperor plugin in QIIME2 [18].
Major taxa with a relative abundance of at least 0.5% in the control and cannulated steers’ rumen microbiota were evaluated. To assess differences in abundance among major taxa between the control and cannulated steers, we used the analysis of composition of microbiomes (ANCOM) plugin implemented in QIIME2 [19]. To analyze the differences between bacterial communities in the control and cannulated steers, we conducted permutational multivariate analysis of variance (PERMANOVA) using PAST3 software [20]. This analysis involved 9999 random permutations. The pH differences between the control and cannulation groups were compared on the preoperative and postoperative days using the Student’s t-test. Additionally, we used orthogonal polynomial contrasts to investigate the longitudinal shifts in major taxa, alpha diversity indices, and pH values from day 0 to day 28 in the control and cannulation groups.
RESULTS AND DISCUSSION
In this study, we defined major taxa, which were primarily focused upon, as those representing at least 0.5% of the total sequences averaged across all 80 samples. Among these taxa, Bacteroidota was identified as the most dominant phylum, accounting for an average of 47.4% of the total sequences across all 80 samples, followed by Firmicutes (31.9%), Verrucomicrobiota (5.2%), Proteobacteria (4.1%), Patescibacteria (3.0%), Spirochaetota (2.2%), Planctomycetota (2.0%), Cyanobacteria (1.5%), Fibrobacterota (0.8%), and Desulfobacterota (0.5%) (Fig. 1A). The dominance of Bacteriodota and Firmicutes observed in this study is consistent with results of previous studies involving Hanwoo steers [21,22]. The ANCOM revealed no significant differences (p > 0.05) in the abundance of major phyla between the control and cannulation groups on the preoperative and postoperative days. Hence, the relative abundance of major phyla might not have been affected by rumen cannulation surgery.
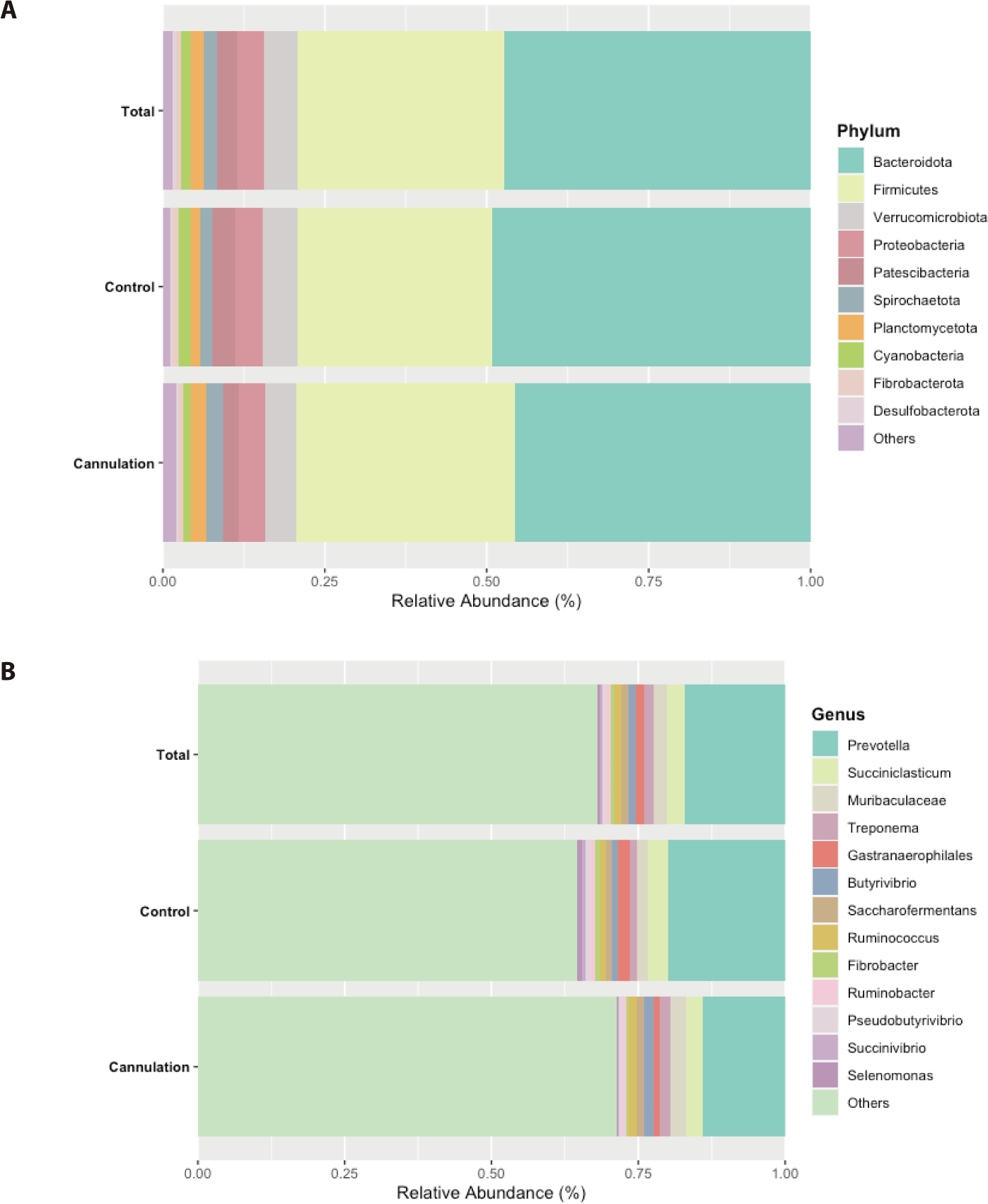
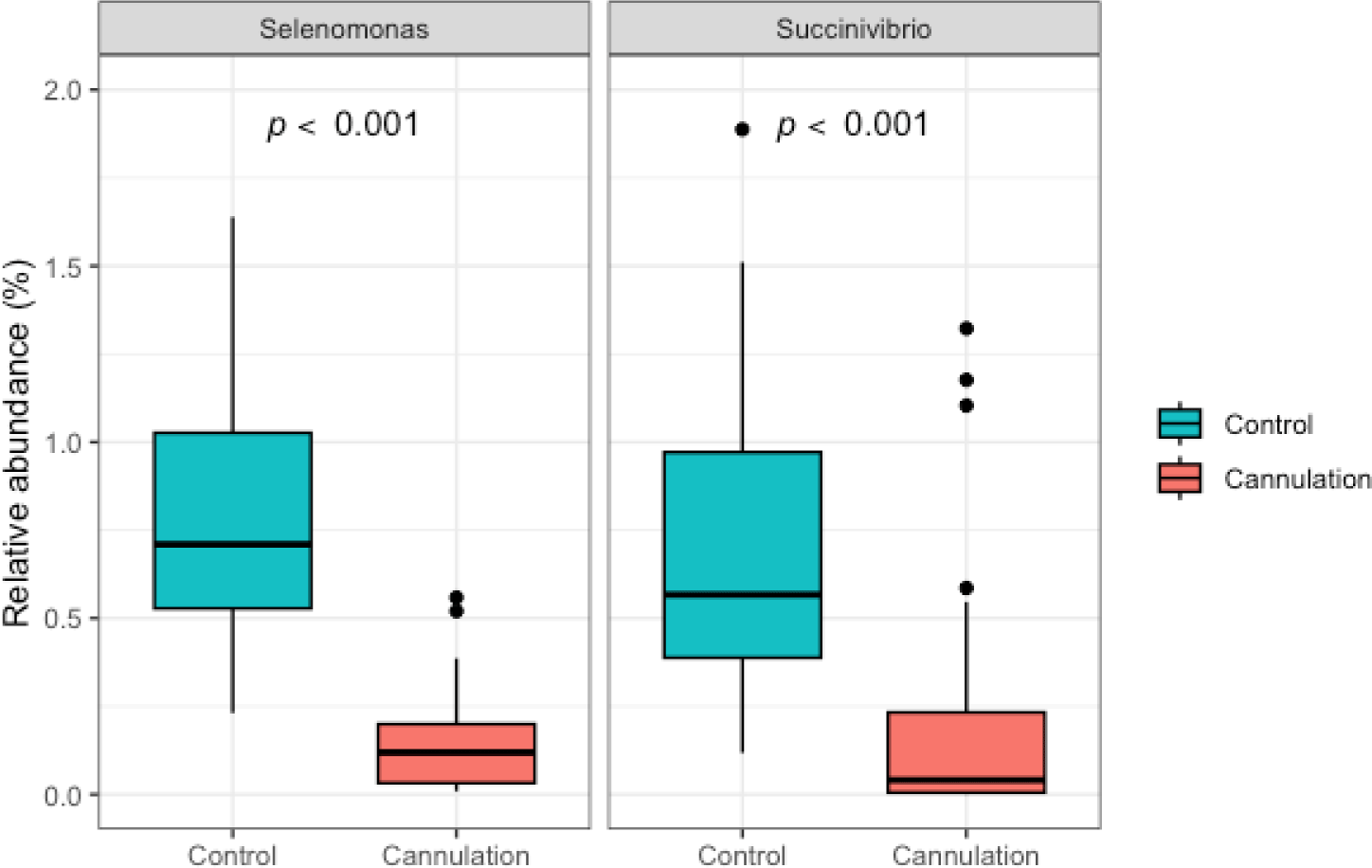
At the genus level, Prevotella was identified as the most dominant, representing 17.0% of the total sequences on average across all 80 samples, followed by Succiniclasticum (3.1%), Treponema (1.6%), Butyrivibrio (1.3%), Saccharofermentans (1.2%), Ruminococcus (1.1%), Fibrobacter (0.8%), Ruminobacter (0.7%), Pseudobutyrivibrio (0.6%), Succinivibrio (0.5%), and Selenomonas (0.5%) (Fig. 1B). The dominance of Prevotella in Hanwoo steers seen in this study conforms to the results of a previous study [21]. On the preoperative day, the ANCOM showed no significant differences (p > 0.05) in the abundance of all major genera between the control and cannulation groups. This result indicates the absence of variances in major genera among individual animals, irrespective of which group they belonged to. However, on the postoperative days, the Selenomonas and Succinivibrio genera were more abundant (p < 0.05) in the control group than in the cannulation group, indicating that the abundance of these two genera might have been affected by cannulation surgery (Fig. 2). Furthermore, the differences in these two genera likely persisted and did not return to their initial states by postoperative day 28. Additional studies are required to confirm if their abundance recovered after 28 days. A previous study showed significant differences among various taxa of goats between the control and cannulation groups [23]. However, this study did not compare rumen microbiota composition between the two groups before cannulation surgery. Therefore, the differences in rumen microbes between the two groups may be attributed to variations among individual animals, regardless of rumen cannulation surgery. Wang et al. [9] reported that the methanogen community was influenced by rumen cannulation. However, they did not compare the methanogen community between the non-cannulated and cannulated groups before the rumen cannulation surgery. Thus, variations among individual animals may result in differences in the methanogen community between the two groups, irrespective of rumen cannulation surgery.
In the control group, both linear and quadratic changes (p < 0.05) were observed in the relative abundance of Butyrivibrio and Selenomonas from day 0 to day 28, while a quadratic change (p < 0.05) was observed in that of Pseudobutyrivibrio from day 0 to day 28 (Table 2). However, in the cannulated group, linear and quadratic changes (p < 0.05) were observed in the relative abundance of Prevotella and Saccharofermentans from day 0 to day 28, while a quadratic change (p < 0.05) was observed in that of Pseudobutyrivibrio from day 0 to day 28 (Table 2). This result indicates that cannulation surgery increases the relative abundance of Prevotella and decreases that of Saccharofermentans. Prevotella is the predominant genus in Hanwoo steers and plays an important role in degradation of various proteins and polysaccharides, including fibers [24,25], while Saccharofermentans is involved in breaking down carbohydrates, including hemicellulose, pectin, and starch [26]. Therefore, the shifts of these two genera due to cannulation surgery may alter rumen fermentation. Further investigations are necessary to determine whether their abundance returns to the original state after day 28. The similarity in quadratic change in relative abundance of Pseudobutyrivibrio in the two groups indicates that it is not likely to be influenced by cannulation surgery.
On the preoperative day, the alpha diversity indices exhibited no significant differences (p > 0.05) between the control and cannulation groups (Table 3). However, on the postoperative days, the alpha diversity indices were significantly greater (p < 0.05) in the control group than in the cannulation group (Table 3). This observation aligns with findings of a previous study involving goats, which revealed that the Chao1 index tended to be higher in the non-cannulated group than in the cannulated group [23]. Although no significant differences were observed between these two groups in another study [9], the alpha diversity indices were numerically greater in the non-cannulated group than in the cannulated group. Hence, microbial diversity may be influenced by rumen cannulation surgery.
Eight rumen samples were collected from eight steers on the preoperative day (day 0) (4 animals × 2 groups × 1 timepoint).
In the control group, Chao1 and Simpson’s diversity indices showed linear changes (p < 0.05), while the alpha diversity indices, except Shannon’s diversity index, demonstrated quadratic changes (p < 0.05) (Table 4). In the cannulation group, however, Shannon and Simpson’s diversity indices showed linear changes (p < 0.05), while all the alpha diversity indices demonstrated quadratic changes (p < 0.05) (Table 4). Shannon’s diversity index greatly decreased from day 4 to day 14 in the cannulation group, indicating that microbial diversity initially decreased after cannulation surgery and then returned to its initial state by postoperative day 28 (Table 4). During the cannulation surgery, the incision made in the rumen seemed to expose a significant number of anaerobic rumen microbes to external oxygen, thereby decreasing their population during the early stages of the procedure. However, as the surgery progressed and the rumen returned to its anaerobic state, the diversity of rumen microbes gradually recovered and returned to its original state.
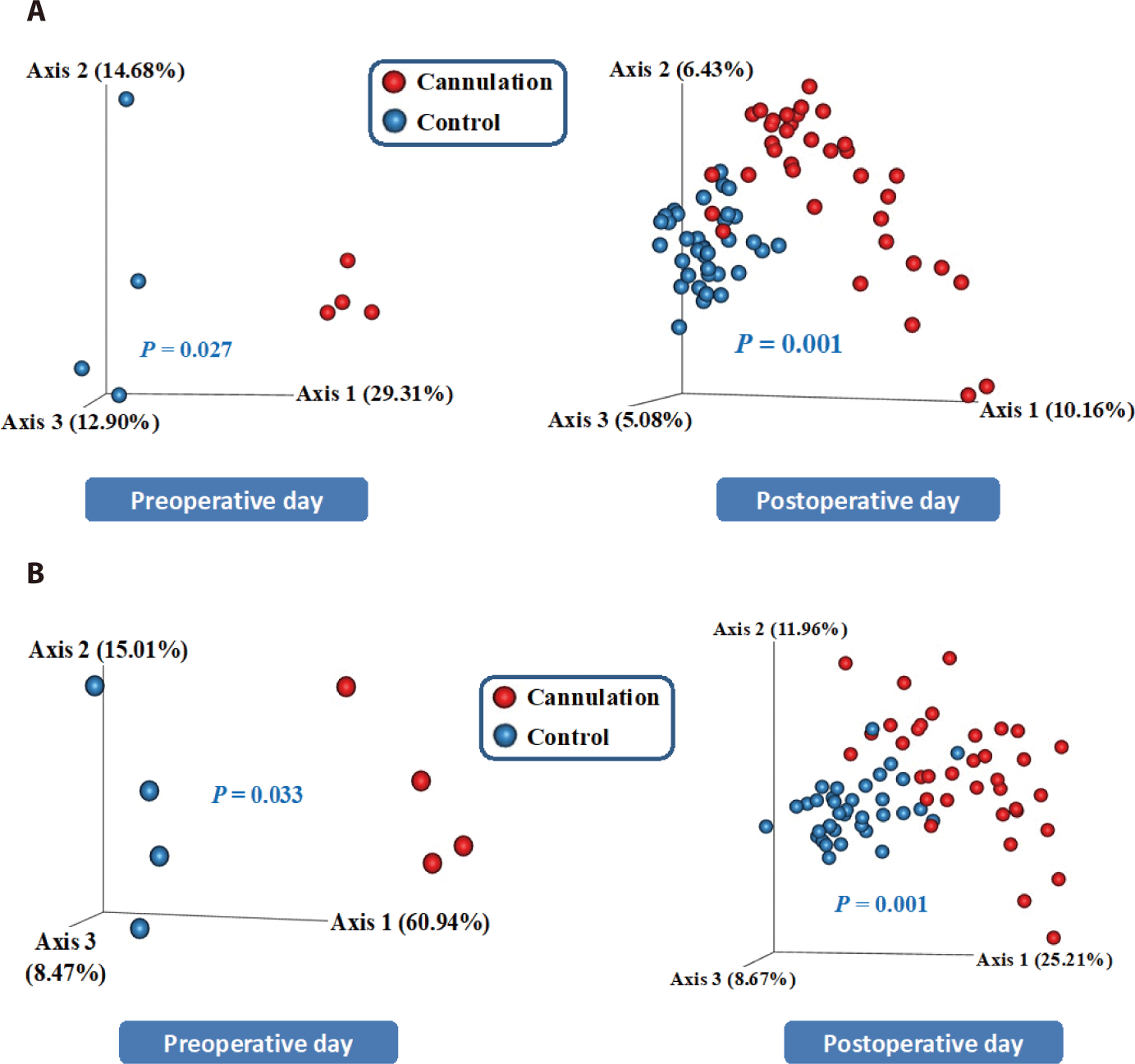
Both the unweighted and weighted UniFrac PCoA plots revealed significant differences (p < 0.05) in the overall microbial composition between the control and cannulation groups, not only on the preoperative day but also on the postoperative days (Fig. 3). On the preoperative day, the ANCOM did not reveal significant differences in the major taxa between the two groups, yet the PCoA plots demonstrated significant differences in the overall microbial composition (Fig. 3A). Therefore, these differences are likely influenced by variations in minor taxa among individual animals before cannulation surgery. On the postoperative day, the ANCOM revealed that only the Selenomonas and Succinivibrio genera exhibited significant differences between the two groups. Therefore, the differences observed in the PCoA plots may be attributed to variations in minor taxa including these two genera between the two groups due to the cannulation surgery. However, these differences might have also been caused by variations in minor taxa among individual animals, regardless of cannulation surgery because the two groups exhibited significant differences before the cannulation surgery.
In the control group, both unweighted and weighted PCoA plots revealed that all pairwise comparisons were not significantly different (p > 0.05), indicating that the overall microbial composition remained unchanged as the days passed (Fig. 4A). Conversely, in the cannulation group, the weighted PCoA plots revealed significant differences (p < 0.05) in pairwise comparisons, indicating differences in the abundance of microbial taxa between the first week and the subsequent days after the cannulation surgery (Fig. 4B). Therefore, the rumen cannulation method can potentially change the composition of rumen microbiota in Hanwoo steers. Further studies are needed to confirm if the composition recovers after 28 days.
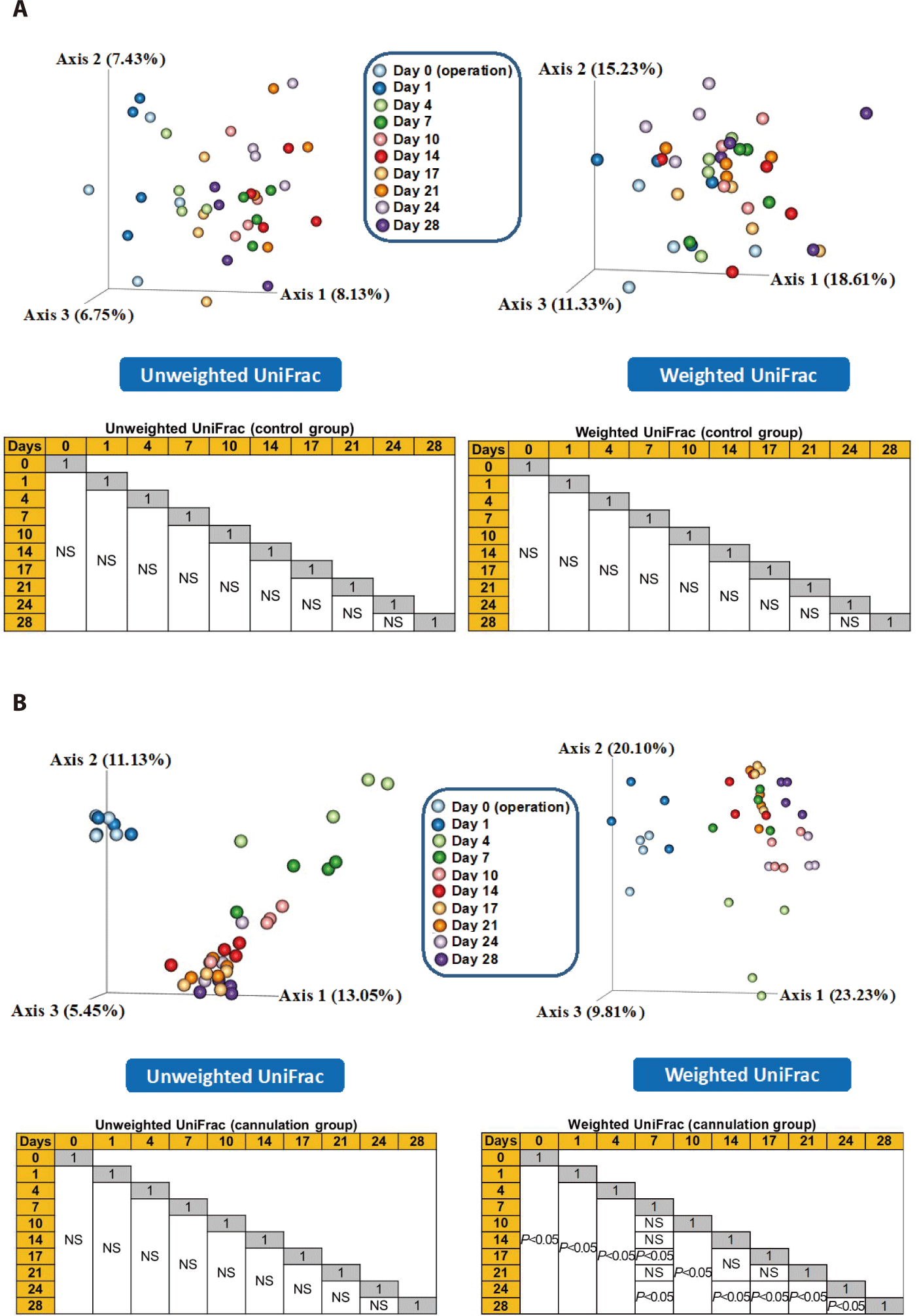
The composition of gut microbiota can undergo shifts after injury due to dysregulation of immune function and inflammation [27]. Therefore, an initial injury resulting from cannulation surgery may also influence the composition of rumen microbiota in Hanwoo steers. Therefore, we conducted a comparative analysis of rumen microbiota between day 0 and days 1 plus 4 to evaluate any potential early-stage effects on the composition of rumen microbiota. The ANCOM showed no significant differences (p > 0.05) in the abundance of all major genera between these two day categories in both the control and cannulation groups. The alpha diversity indices did not differ (p > 0.05) between the two day categories in the control group, while Chao1 was greater (p < 0.05) in the day 0 category than in the days 1 plus 4 category in the cannulation group (Table 5). Although no significant differences were observed in Shannon and Simpson’s diversity indices, they were numerically greater in the day 0 category than in the days 1 plus 4 category in the cannulation group. In the early stages of cannulation surgery, microbial diversity might have reduced owing to the injury caused by the procedure. Although the major genera did not differ between these two day categories in the cannulation group, the minor genera might have shifted, as indicated in the PCoA plots (Fig. 4). This alteration in microbial diversity may be attributed to the exposure of a significant number of anaerobic rumen microbes to external oxygen during the early stages of the procedure.
On the preoperative day, ruminal pH did not exhibit significant differences between the control and cannulation groups (Table 6). However, on the postoperative days, significant differences (p < 0.05) in ruminal pH were observed between the two groups (Table 6). Wang et al. [9] also showed that ruminal pH exhibits significant differences between the control and cannulation groups. No linear or quadratic changes were observed in either the control or cannulation group from day 0 to day 28 (Table 7). However, ruminal pH increased due to the rumen cannulation surgery and subsequently returned to its initial state by postoperative day 28 (Table 7). These changes may be associated with alterations in microbial diversity due to exposure of anaerobic rumen microbes to external oxygen during the early stages of the procedure.
| Treatment | Preoperative day1) | Postoperative days2) |
|---|---|---|
| Control | 6.719 (n = 4) | 6.646 (n = 36) |
| Cannulation | 6.913 (n = 4) | 7.066 (n = 36) |
| SEM | 0.130 | 0.046 |
| p-value | 0.500 | < 0.001 |
CONCLUSION
The study findings indicate that rumen cannulation surgery can potentially influence the composition of rumen microbiota in Hanwoo cattle. However, the major genera, microbial diversity, and pH levels appeared to gradually return to their original state over approximately 1 month. The current data show that the composition of predominant rumen microbes is affected by rumen cannulation surgery. Additionally, minor rumen microbes that are altered by this procedure do not return to their initial states by postoperative day 28.