













INTRODUCTION
The demand for alternative feed ingredients is increasing due to unstable costs of universal feed ingredients, such as corn and wheat, caused by unstable international situations such as continuous climate change and wars. In those circumstance, feed formulators should have considered the various alternatives which has low nutrient value or has high price. Thus, we were focused on Italian ryegrass (IRG) as an alternative feed ingredient for pig. Ryegrass has two major species, Lolium perenne, also called English ryegrass and Lolium multiflorum Lam., also called IRG. IRG has good characteristics which high-yielding, rapid seedling establishment, weed suppression, and palatable forage that is tolerant to various environmental conditions [1,2]. Even in 2004, a new cold-tolerant and early-heading variety of IRG called “Kogreen” was developed to be suitable in Korean seasonal conditions [3]. Generally IRG regarded as good ingredient for ruminant without affecting negative effect on the animal. Recent research also suggests that Hanwoo fed IRG improves carcass yield 12.5% higher than control diet [4]. However, there is limited research on IRG diets for non-ruminant animals due to their high fibers. Despite the characteristics of fibers in monogastric animal diet which generally regarded as an anti-nutritional actor because of its lower protein level and energy digestibility, fibers could improve gut health with by-product from bacteria which increasing gut length, mass, and villus height [5]. Non-starch polysaccharide (NSP), some portion of dietary fibers, in diet is also generally considered as anti-nutritional factors for non-ruminant animals because of their characteristics which encapsulate nutrients intracellularly and cannot be degraded by endogenous enzymes of non-ruminant animals [6]. NSPs also have cation exchange capacity, hydration properties, viscosity, and organic compound absorptive properties at animal digestive tract [7]. Because of the characteristics of NSP, Choct, Dersjant-Li [8] suggests that NSP in diet, especially for young monogastric animals, NSP decrease the nutrient digestibility, proliferation of Escherichia coli causing swine dysentery and dehydration of NSP could reduce negative effects of NSP. Therefore, previous dietary NSP studies focused on mitigating the effects of NSPs by adding NSP enzymes, and this addition is suggested to have expected and unexpected health benefits for swine in NSP diets [9]. Following studies, Li et al. [10] suggests weaning pig fed high barley contained diet with NSP enzymes could improves growth performance and increase gamma GT activity in jejunal mucosa and Ao et al. [11] suggests growing pig fed corn-soy based diet with multi NSP enzymes improve average daily gain (ADG), G:F, and improve digestibility of dry matter, nitrogen and amino acids. Besides, the microbiome is also crucial for its host animals, as it can provide various benefits that the host lacks due to its genetic diversity [12]. Animals are born without microbial contamination; however, they are continually exposed to microbes due to their gastrointestinal structure, which resembles an open tube [13]. Therefore, feed ingredients consumed by animals are important for shaping the gut microbial community. However, research of IRG in the monogastric animal diet is very limited. Research by Recharla et al. [14] suggests that IRG, with exogenous enzyme addition, has no impact on growth performance and nutrient digestibility; however, it affects the microbial composition in the hindgut. Park et al. [15] propose that adding a maximum of 1.5% IRG to swine diets can reduce odorous compounds in manure by altering the microbial community, while maintaining consistent growth performance.
Therefore, this study was conducted to estimate the availability, tolerance level, and effects of IRG with NSP enzyme supplementation on growth performance, gut microbiome, blood parameter, and manure odor emission in finisher pigs.
MATERIALS AND METHODS
Following AOAC methods [16], samples were initially pre-dried at 60°C in a drying oven and ground. The feed ingredient was assessed for gross energy using a bomb calorimeter (Model C2000, IKA, Staufen, Germany), ether extract, crude fiber (CF), ash, neutral detergent fiber (NDF), and acid detergent fiber (ADF). Calcium was determined using an atomic absorption spectrophotometer (Perkin Elmer 3300, Perkin Elmer, Waltham, MA, USA). Phosphorus content in the feed ingredients, experimental diets, and feces was measured using a spectrophotometer (Optizen 2120 UV, Mecasys, Daejeon, Korea). The calculated composition of IRG was as follows: 1,447.25 kcal/kg metabolizable energy, 12.1% crude protein, 3.6% ether extract, 22.3% crude fiber, 0.2% phosphate, and 0.4% calcium.
Six experimental diets were formulated and fed to the animals. The diets consisted of two enzyme levels (non and recommended) and three dietary IRG levels (0%, 2.5%, and 5%). All experimental diets were formulated to match equal nutrient levels of metabolizable energy (ME) and crude protein (CP). The formulation and calculated values of experimental diets are presented in Table 1. In total, 60 crossbred Landrace × Yorkshire × Duroc (LYD) pigs were used to assess the effects of dietary IRG and multi-enzyme addition. Multi-enzymes were added to the diet to achieve the commercially recommended level (255 U/kg cellulase; 1,250 U/kg beta-mannanase; 6,000 U/kg xylanase). The pigs were randomly assigned to six dietary treatments (two enzyme levels × three IRG levels), with each treatment replicated five times, and each pen containing two pigs. The average body weight (BW) of the allocated pigs was 88.35 ± 2.57 kg. All pigs are allotted with completely randomized block design. The experiment lasted for four weeks, during which the experimental diet and fresh water were provided ad libitum via metal troughs and nipple drinkers.
1) Vitamin-Mineral premix provided the following nutrients per kg: Vitamin A, 15,000 IU; Vitamin D3, 2850 IU; Vitamin E, 75 IU; Vitamin K3, 4.5 mg; Vitamin B1, 3.375 mg; Vitamin B2, 7.5 mg; Vitamin B6, 4.875 mg; Vitamin B12, 56.250 mg; Calcium D-Pantothenate, 25.5; Endox, 18 mg; Niacin, 48 mg; Folic acid, 1.875; Iodine, 0.6 mg; Iron, 150 mg; Magnesium, 54 mg; Zinc, 90 mg; Copper, 39 mg; Biotin, 0.45 mg.
Growth performance data were collected to determine the effects of IRG and multi-enzymes present in the diet on finisher pig. All measurements were taken on a pen basis. Initial and final body weights were recorded on the first and last days of the experiment. Average daily feed intake (ADFI) was determined by measuring the total feed consumption. ADG and feed efficiency (FE) were calculated using the measured body weights and feed intake.
Fecal samples were analyzed to estimate the effects of IRG and multi-enzymes on fecal microbiome and odorous compounds. Fecal samples were collected from the anus after rectal massage and stored in a −80°C deep fridge with 50-mL conical tube.
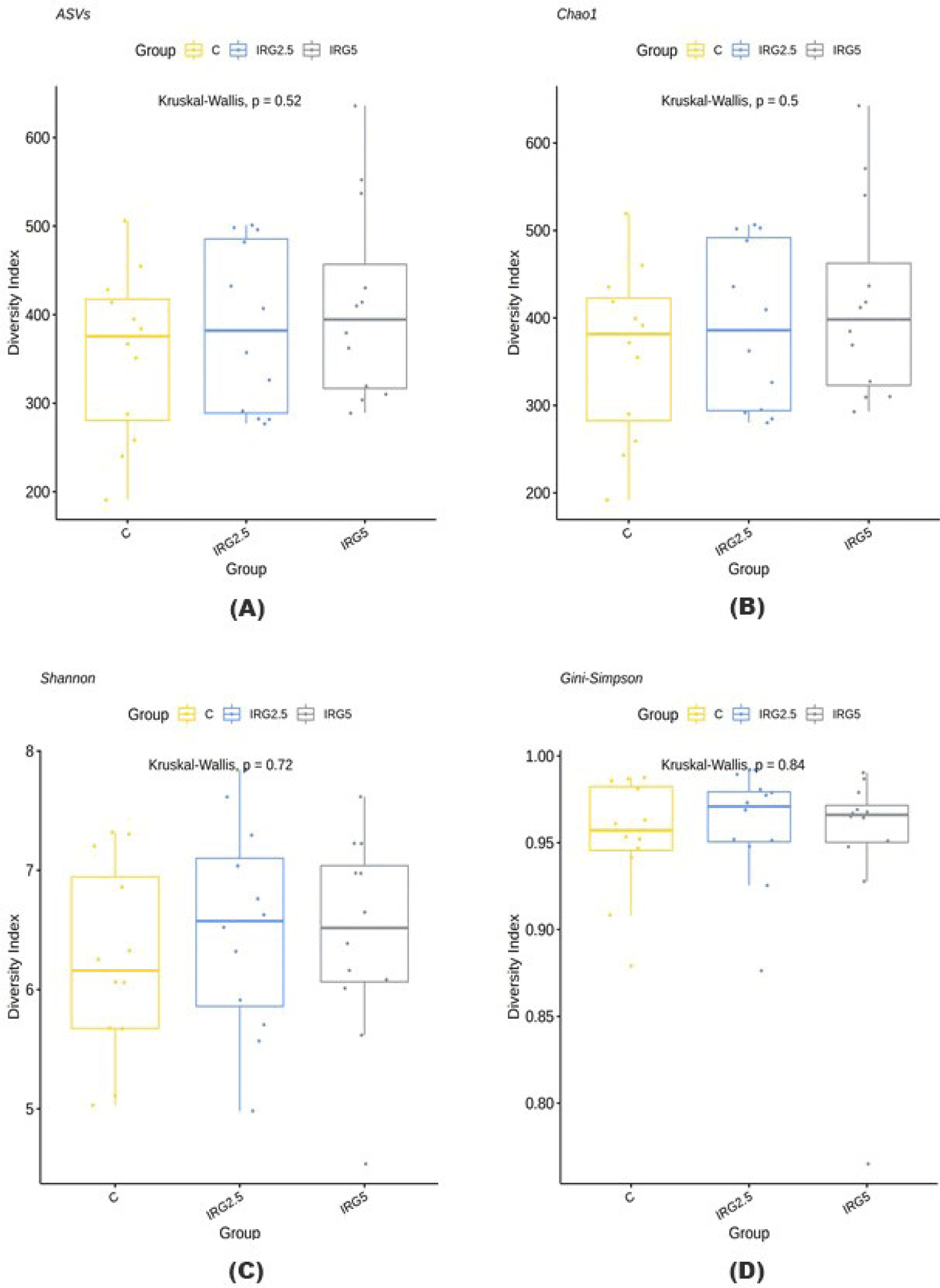
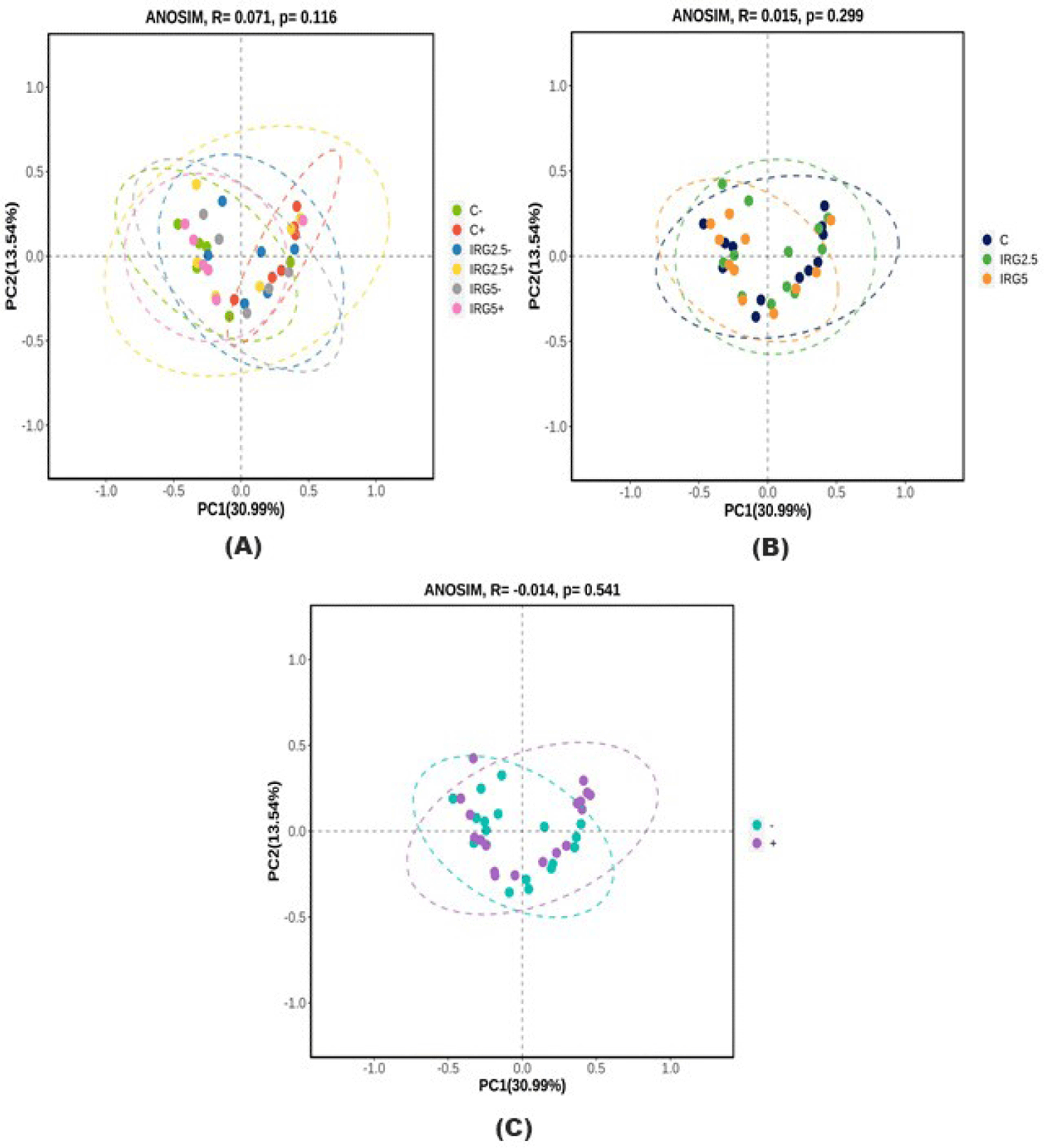
A G-spinTM genomic DNA extraction kit was used to extract genomic DNA from fecal samples. The processed DNA was analyzed using 16S rRNA [17]. All processed sequences were clustered at a 97% similarity threshold using QIIME (v.1.8.0). The alpha diversity of the microbiome at the IRG level was assessed by calculating amplicon sequence variants (ASV), Chao1, Shannon, and Gini-Simpson diversity indices, whereas beta diversity was evaluated using the Bray–Curtis distance, and principal coordinate analysis (PCoA) was employed to visualize it.
Fecal volatile fatty acids, phenols, and indoles were analyzed following the procedure described by Flickinger et al. [18]. Volatile fatty acids were determined using a gas chromatograph (GC, 6890N, Agilent, Santa Clara, CA, USA). Phenols and indoles were extracted by mixing 2 g of feces with 5 mL of methanol containing 2,000 ppm of 5-chloroindole (internal standard). The feces–methanol mixture was sealed with parafilm, thoroughly mixed, and incubated for 1 h at 4°C with frequent mixing. Thereafter, the tubes were centrifuged at 29,000×g for 20 min at 4°C, and the supernatant was collected. The remaining pellet was mixed again with 5 mL of methanol and extracted as described above. The two supernatant fractions were combined for gas-liquid chromatography analysis. The individual concentrations of indole, phenol, p-cresol, and 4-ethylphenol were determined using a gas chromatograph (GC, 6890N, Agilent).
All collected and calculated data were analyzed using two-way analysis of variance (ANOVA) in SPSS 26.0 (SPSS, Chicago, IL, USA). The pen and individual pig were considered the statistical unit for growth performance analysis and manure analysis, respectively. A p < 0.05 was considered statistically significant. In cases where significant differences were found, Tukey’s post hoc test was employed to examine the differences between the groups.
RESULTS AND DISCUSSION
Table 2 shows the impact of dietary IRG and multi-enzymes on growth performance. Initial BW, final BW, ADG, and FE showed no significant differences in terms of p-value, IRG, enzyme, or interaction effects. However, ADFI had a significant difference (p < 0.05), with enzymes influencing it. Exogenous carbohydrase enzymes are considered enhancers, improving energy availability and nutrient digestibility by breaking down indigestible bonds in swine diets. They may also enhance intestinal microbiomes, potentially improving host health [19–21]. Previous research suggests that adding NSP-degrading enzymes to swine diets can boost feed intake [22,23]. Combining these enzymes with others can enhance swine diet performance more effectively than using only one type of enzyme [24]. Our results align with previous studies indicating that exogenous NSP-degrading enzymes increase feed intake. However, no significant differences in any growth performance parameters were observed until IRG was added at a 5% level. In general, ingredients with higher fiber content are considered to have lower nutritional value for non-ruminants because fibers cannot be broken down by the host animal’s endogenous enzymes [25]. Previous studies also agreed that a high fiber diet can negatively affect the growth performance of pigs at various growth stages [26–29]. However, the effect of dietary fiber on growth performance remains controversial. Some research suggests that high fiber diets decreases growth performance in growing pigs but have a limited effect on finisher pigs [30,31], whereas other studies propose that high fiber diets have no impact on growing pigs [32,33]. There are very limited results of IRG in swine diet; however, these suggest that a small amount of IRG does not affect the growth performance [14,15]. Agyekum et al. [29] also suggests that the conflicting results of high fiber effects on swine does not depend only on the fiber amount, such as CF, NDF, ADF, and NSP, but on the ratio between the fiber components. The absence of interaction effects between IRG and enzymes may be due to the limited role of 5% IRG as an anti-nutritional factor in the diet.
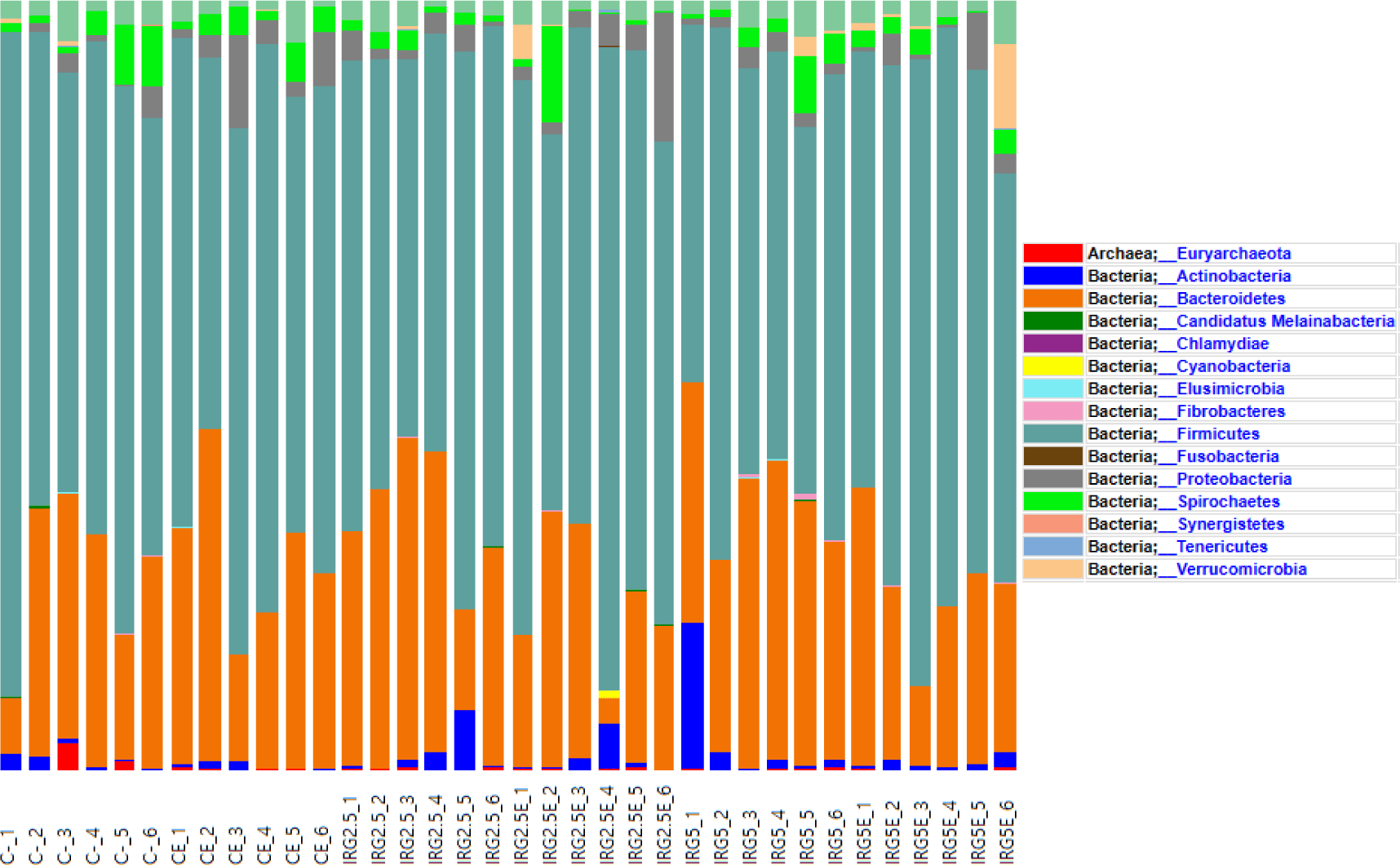
Taxonomic assignment at the phylum level, alpha diversity, and beta diversity are analyzed and illustrated in Figs. 1, 2, and 3 to check the effect of dietary fibers on the gut microbiome. According to Kim et al. [34], the majority of bacteria can be classified into five phyla: Firmicutes, Bacteroidetes, Proteobacteria, Actinobacteria, and Spirochaetes, with Firmicutes and Bacteroidetes making up 90% of the bacterial population. These phyla are known to interact with resistant starch [35], and Bacteroidetes possess high carbohydrate enzyme gene activity for breaking down plant cell wall components like glucuronoxylans, xyloglucans, and pectin [36]. As reported by Patil et al. [37], the addition of dietary NSP stimulates the commensal microbiome to produce short-chain organic acids, lowering the gut pH. Consequently, we aimed to investigate potential changes in microbiome diversity with the addition of dietary IRG. Alpha diversity analysis employed ASVs, Chao1, Shannon, and Gini-Simpson methods. However, our results did not indicate any significant differences in alpha diversity. Similarly, beta diversity showed no significant distinctions between the groups. Agyekum and Nyachoti [5] suggested about the ambivalence of the dietary fiber in swine gut which even in the same soluble fiber research which some are suggests that soluble fiber in swine diet contribute the risk of colibacillosis and swine dysentery by colonized pathogenic bacteria caused by the low viscosity but the others research are not. Zhang et al. [38] also suggested that NSP enzyme addition did not impact alpha diversity in growing pigs. Consequently, the results of this study suggested that 5% IRG in the diet with NSP enzymes can be managed without negatively altering microbial composition. The results of the analysis of fecal odorous compounds are shown in Table 3. Fecal odorous compounds analysis was decided because of believes that the dietary NSP could affect the gut microbiome. The major odorous compounds in swine manure are volatile organic compounds, indoles, and phenols [39–41]. Furthermore, whether microbiomes affected by substrates in diet, the major end-products in manure are acetate, propionate, and butyrate [42]. Based on a previous research, volatile fatty acids produced by Eubacterium and Clostridium are the major contributor to odorous compounds. However, various studies presented that the gut microbiome is affected by dietary ingredients, especially fibers [43–45]. Additionally, the gut microbial metabolism of tryptophan results in the production of indole compounds [46]. Therefore, IRG with additional multi-enzymes could possibly change the odorous compounds by reducing the precursor of volatile organic compounds, but no significant difference was observed, except for the indole level influenced by the enzyme. We speculate that this result arises from the degradation of the fiber structure, leading to the release and utilization of entrapped nutrients. From those results, we suggest that adding 5% IRG in finisher pig diets can help achieve the desired goals without negatively impacting the growth performance. Also adding enzymes in high fiber diet can also enhance feed intake and reduce indole compounds by degrading the fiber source.
In conclusion, 5% IRG with multi NSP enzymes could be an option for replacing corn in swine diet and following research which set the maximum IRG level without affecting growth performance and benefits of replacing IRG to corn by economic analyzing are required.