RESEARCH ARTICLE
Evaluation of zinc oxide and copper oxide nanoparticles as potential alternatives to antibiotics for managing fowl typhoid in broilers
Muhammad Atif Raza1,#
,
Eungyung Kim1,#,
Muhammad Shakeel1,2,
Muhammad Fiaz2,
Lei Ma1,
Hyeonjin Kim1,
Chae Yeon Kim1,
Zhibin Liu1,
Ke Huang1,
Kanghyun Park1,
Muhammad Tariq Javed3,
Myoung Ok Kim1,*
Author Information & Copyright ▼
1Department of Animal Science and Biotechnology, Research Center for Horse Industry, Kyungpook National University, Sangju 37224, Korea
2Faculty of Veterinary and Animal Sciences, Pir Mehr Ali Shah, Arid Agriculture University, Rawalpindi 44000, Pakistan
3Department of Pathology, Faculty of Veterinary Science, University of Agriculture, Faisalabad 38000, Pakistan
#These authors contributed equally to this work.
*Corresponding author: Myoung Ok Kim, Department of Animal Science and Biotechnology, Research Center for Horse Industry, Kyungpook National University, Sangju 37224, Korea. Tel: +82-54-530-1234, E-mail:
ok4325@knu.ac.kr
© Copyright 2024 Korean Society of Animal Science and Technology. This is an Open-Access article distributed under the terms of the
Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits
unrestricted non-commercial use, distribution, and reproduction in any
medium, provided the original work is properly cited.
Received: Jun 09, 2023; Revised: Aug 05, 2023; Accepted: Aug 31, 2023
Published Online: Sep 30, 2024
Abstract
Antimicrobial resistance poses challenges to humans and animals, especially to the poultry sector in control of fowl typhoid with antibiotics, leading to increased mortality and food insecurity. Therefore, it is essential to develop more effective medications as alternatives to antibiotics. Currently, zinc oxide and copper oxide nanoparticles are of such significant interest due to their antibacterial properties. This study aimed to evaluate antimicrobial activity of zinc oxide and copper oxide nanoparticles against fowl typhoid in broilers. Ninety broiler chicks were raised under suitable management conditions. On day 10 of age, chicks were divided into six groups: control negative, control positive, T1, T2, T3, and T4. On day 19 of age, chicks in all groups except control negative were infected with Salmonella gallinarum (0.2 mL, 108 CFU/mL). After appearance of clinical signs, the treatments (Florfenicol; 50 mg/L drinking water [T1], and zinc oxide + copper oxide nanoparticles; 25 + 10 mg/kg/d [T2], 37.5 + 15 mg/kg/d [T3], and 50 + 20 mg/kg/d [T4]) were administered to chicks. Chicks were sacrificed on 26th and 30th day of age, and samples of blood and tissue were obtained. Hematological analysis with gross and histopathological examination of spleen, thymus and bursa of Fabricius was performed. Results revealed that there was no visible congestion in spleen and thymus of T3 and T4 at 11th day post infection. Antibody level against new castle’s disease and lymphoproliferative response showed no significant difference in all groups. However, phagocytic response in nanoparticles treated groups exhibited a notable (p < 0.01) distinction compared to control positive. Notably, T3 demonstrated the highest level of phagocytic activity. Hematological parameters, including lymphocytes, heterophils, eosinophils, and heterophils/lymphocytes ratio in groups T2, T3, and T4, indicated significant (p < 0.01) difference compared to control positive. However, lymphocytes, heterophils, and heterophils/lymphocytes ratio in groups T2, T3, and T4 showed no significant difference when compared to T1. Nanoparticle treated groups showed decreased (p < 0.01) congestion of spleen and thymus as compared to control positive. Overall, zinc oxide and copper oxide nanoparticles have potential to serve as an alternative to florfenicol in treatment of fowl typhoid.
Keywords: Antimicrobial resistance; Salmonella gallinarum; Infection; Poultry health; Immunology
INTRODUCTION
The poultry sector has emerged as a vital industry in growing economies [1]. It not only meets the daily protein requirements of the growing population but also provides opportunities for employment [2] through the supply of high-quality food items such as chicken meat and eggs, ensuring global food security and nutrition [3,4]. Poultry meat and eggs are important for human health because the protein and vitamins they contain play a crucial role in the development of immunity [5].
However, the poultry industry faces several challenges that jeopardize economic output and the health of animals and humans in many countries [6]. Poultry mortality is one of the main issues that hinder continuous food supply to the population and high mortality may be largely attributed to the spread of infectious diseases [7]. Fowl typhoid is a septicemic disease of poultry caused by gram negative bacterium Salmonella gallinarum which produces the endotoxins in the blood circulation of host. The disease is associated with substantial losses to the country’s economy through high mortality and decreased egg production [8]. Infectious diseases like salmonellosis, new castle’s disease (NDV), infectious bursal disease etc. are highly occurring diseases in poultry around the world causing high mortality in the birds [9]. These diseases are associated with the immunosuppression of the affected birds by damaging the immune organs leading to mortality [10]. The birds affected by fowl typhoid manifest clinical signs like depression, pale and shrunken comb, ruffled feathers, anorexia, dyspnea, huddling, diarrhea, and adherence of the excreta to the vent [8], and inflammation in the liver, spleen, cecum, and yolk sac [11]. The morbidity rate is very high, resulting in 93%–100% mortality occurring when the birds are infected with a bacterial load of 108 colony forming unit per milli liter (CFU/mL) [12]. Therefore, it is necessary to reduce the mortality in poultry from infectious diseases.
Antibiotics are widely used for the prevention and treatment of various infectious diseases as well as growth promoters in poultry [13]. Although antimicrobial agents play a vital role in the control of morbidity and mortality in animals as well as humans, the extensive use of these agents has led to the generation of antimicrobial resistance (AMR) in pathogenic bacteria [14]. The irrational or irresponsible use of different antibiotics, especially florfenicol, promotes the prevalence of AMR in poultry farms [15]. The emergence and transmission of AMR strains of different bacteria not only affects poultry production but also threatens human health [16]. The AMR salmonella species can be transferred to humans while handling or slaughtering the infected birds which leads to human illness [17]. Therefore, the presence of AMR in animals raised for food is a significant concern [18] as it poses a substantial zoonotic risk to human health. This is especially true considering the abundance of AMR bacteria such as Salmonella, Campylobacter, and Listeria [19]. Hence, there is an urgent requirement to develop alternative therapeutic treatments that can replace antibiotics.
Nanotechnology is a new field of science with extensive applications in the development of nanomedicine [20]. Several metals including zinc oxide (ZnO) and copper oxide (CuO) nanoparticles have excellent antibacterial activity against gram-positive and gram-negative bacteria [21]. Hameed et al. [22] reported that ZnO nanoparticles can inhibit the growth of Escherichia coli and Klebsiellapneumoniae on the culture plates and increased the zone of inhibition. Similarly, the in vitro antibacterial activity of ZnO nanoparticles against E. coli, Enterobacter aerogenes, Micrococcus luteus, and Bacillus subtilis were also documented [23]. Recently, the in vitro antibacterial activity of ZnO and CuO nanoparticles has been studied against B. subtilis, E. coli, Staphylococcus aureus, Salmonella typhimurium, and Pseudomonas aeruginosa [24]. Kim et al. [25] reported that pigs treated with nano ZnO showed increased average daily weight gain and decreased incidence of fecal score and diarrhoea. Nano ZnO also inhibited the colonization of E. coli, S. typhimurium, and Listeria monocytogenes.
The gram-positive and gram-negative bacteria have different structures of cell wall. The gram-positive bacteria have a thick layer of peptidoglycan in the cell wall while the gram-negative bacteria have thin layer of peptidoglycan and an additional layer of lipopolysaccharide molecules in the cell wall which carry a negative charge. The negative charge have more affinity for positive ions released from nanoparticles, causing an increased uptake of ions leading to intracellular damage in bacterial cell [26]. The antibacterial activity of ZnO and CuO nanoparticles is due to the generation of free radicals and reactive oxygen species that bind to the bacterial cell wall and cause bacterial cell destruction [27]. The CuO nanoparticles are extremely reactive because of their high surface area to volume ratio which improves their antimicrobial efficiency [6]. The metal oxides show antibacterial properties by generating reactive oxygen species and free radicals. The oxygen reacts with the CuO and forms cupric ion (Cu2+); the cation reacts with superoxide ion (O2−), leading to oxidative stress. The O2− ion reduces the Cu2+ ion to cuprous ion and produces hydrogen peroxide (H2O2) which reacts with copper and again produces hydroxyl ion. Similarly, ZnO nanoparticles also produce H2O2 and O2− ion. The H2O2 penetrates the bacterial cells and causes cellular membrane damage, lipid peroxidation, and ultimately bacterial cell growth inhibition and bacterial cell destruction by damaging the cellular components such as deoxyribose nucleic acid and proteins [26,28,29]. Supplementation of nano copper to the poultry diet can improve the daily weight gain, erythrocyte count, and haematocrit level in chicken [30]. Recently, Kim et al. [31] reported that copper is frequently used as growth promoter in monogastric animals. Copper can shift intestinal microbiota in pigs which may be attributed with its antimicrobial activities [32]. It is anticipated that nanoparticles will become the most appropriate antibacterial drugs in the future. Therefore, this study was designed to determine the antibacterial effects of ZnO and CuO nanoparticles in S. gallinarum induced infection in broilers in terms of their hematological, pathological, and immunological parameters.
MATERIALS AND METHODS
Ethics approval and consent to participate
The animal care and experimental protocols used in the present study were approved by the Graduate Study Research Board, in accordance with the guidelines of Institution of Animal Care and Use Committee, University of Agriculture Faisalabad, Pakistan (approval number: DGS/.7049-52).
Experimental birds and study plan
A total of 90, one-day-old broiler chicks (Hubbard) were selected for this study. The birds were kept under the same environment and management conditions for the first 18 days. The ZnO and CuO nanoparticles were prepared at the Department of Physics, University of Agriculture Faisalabad (Pakistan). The chemicals cupric chloride (CuCl2), zinc sulphate (ZnSO4) and sodium hydroxide (NaOH) were kindly provided by Dr. Muhammad Yasir Javed (Department of Physics, University of Agriculture Faisalabad, Pakistan) for the preparation of ZnO and CuO nanoparticles. The CuO nanoparticles were prepared by co-precipitation method as previously documented by Phiwdang et al. [33] using CuCl2 and NaOH as precursor. Briefly, CuCl2 (1 M) was dissolved in distilled water (1 L) and constantly stirred at magnetic stirrer until completion dissolution of CuCl2. After, NaOH (1 M) was added gently drop by drop under vigorous stirring on magnetic stirrer. The black precipitates of CuO were obtained and washed with distilled water several times. Later, the washed precipitates were dried in oven at 80°C overnight and dried product was kept in muffle furnace (500°C) for 4 hours. Finally, CuO was crushed into fine powder. The size and purity of CuO nanoparticles used in the present study were 33.20 nm and 99.9% respectively, as the nanoparticles were from same batch already reported by our research group [6]. The ZnO nanoparticles were prepared by co-precipitation method as previously documented by D et al. [34] using ZnSO4 and NaOH as precursor. Briefly, ZnSO4 (1 M) was dissolved in distilled water (1 L) and constantly stirred at magnetic stirrer for 1 hour. After complete dissolution of ZnSO4, NaOH (2 M) solution was added drop by drop under continuous stirring conditions for two hours. Subsequently, a white creamy suspension was formed and was allowed to settle overnight. The precipitate was several times with distilled water and dried in the oven at 80°C. During drying, zinc hydroxide is completely converted into ZnO. The ZnO was kept in the muffle furnace (500°C) for 4 hours. Finally, ZnO was crushed into fine powder. The size and purity of ZnO nanoparticles used in the present study were 97.5 nm and 99.9% respectively, as the nanoparticles were from same batch already documented by Bahadur [35].
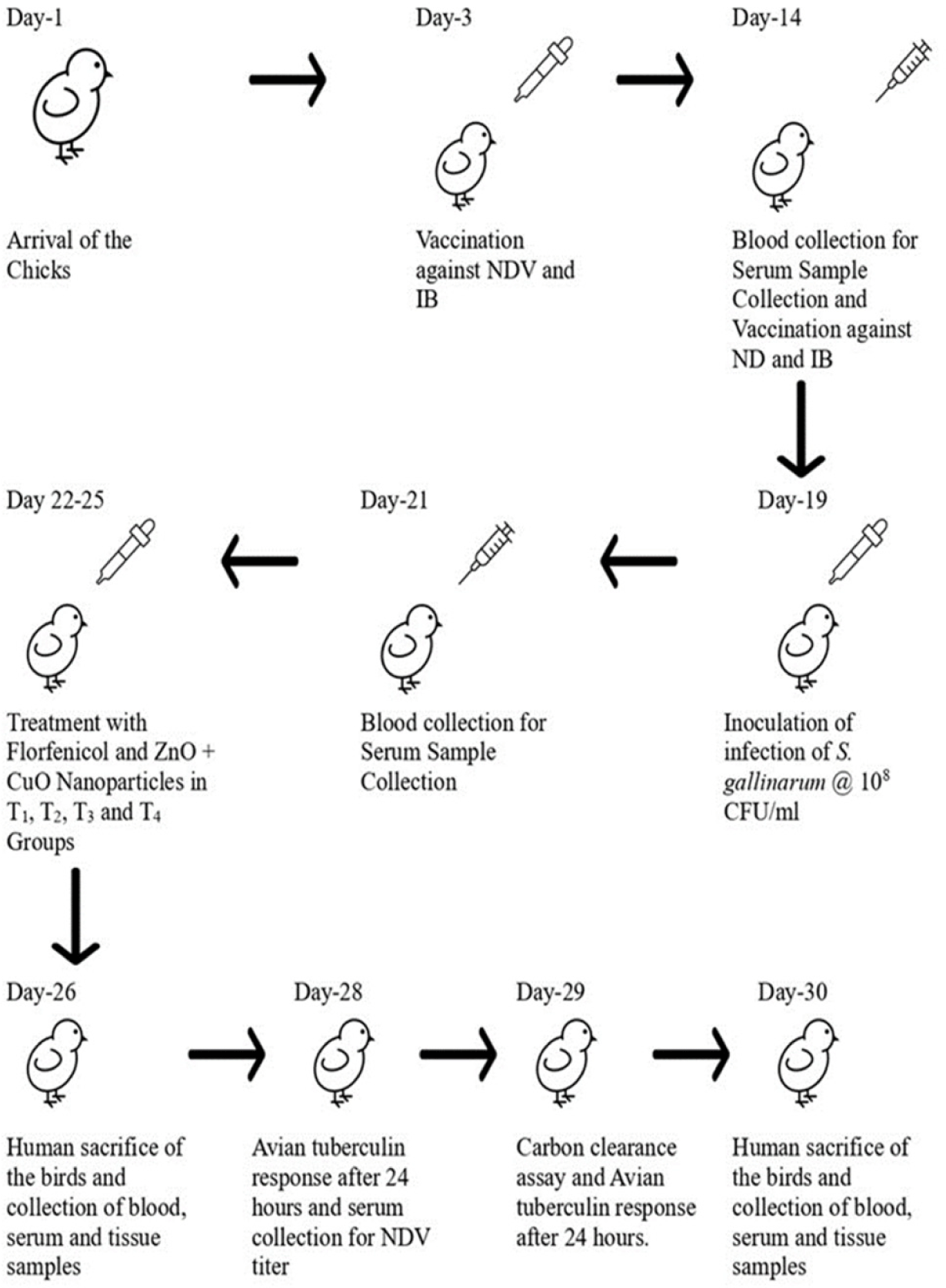
The birds were divided (15 birds/group) into six groups: control negative, control positive, T1 (Florfenicol; 50 mg/L drinking water), T2 (ZnO nanoparticles; 25 mg/kg + CuO nanoparticles; 10 mg/kg), T3 (ZnO nanoparticles; 37.5 mg/kg + CuO nanoparticles; 15 mg/kg), and T4 (ZnO nanoparticles; 50 mg/kg + CuO nanoparticles; 20 mg/kg). The birds were maintained in six individual compartments with wood shavings as litter material. On day 19, the birds of all the groups except the control negative were orally infected with S. gallinarum at a dose of 108 CFU/mL as shown in the experimental design (Fig. 1). All birds were provided with clean water and commercially available feed ad libitum throughout the study. The treatments were given to the birds three days post-infection (after the appearance of clinical signs).
Parameters and data collection
The birds were sacrificed on day 26 and 30 of the study. The blood samples were collected in ethylene diamine tetra acetic acid vacutainers (LOT: 07072014, Lab Vac, Bayswater North VIC, Australia).
Gross pathology and histopathology of spleen, thymus, bursa of Fabricius
Spleen, thymus, and bursa of Fabricius were isolated after sacrifice and inspected for abnormal morphology changes. The scoring of the congestion was performed using an arbitrary scoring system. The congestion was described as none (−), mild (+), moderate (++), or severe (+++).
The tissue samples were taken spleen, thymus, and bursa of Fabricius, cut into small pieces of 2–3 cm with a thickness of 1–2 mm, and placed in containers with 10% neutral buffered formalin solution for fixation, followed by histopathological examination. The tissue samples were dipped in a series of ethanol solutions with different concentrations. After the tissue samples were cleaned with xylene-I and xylene-II to remove the dehydrating agent. Finally, the tissue section slides were prepared, and staining was done. The previously described protocols [36] were used for the processing of the tissue sections and staining with hematoxylin and eosin stains. The quantitative analysis of the histopathological slides was analyzed by using QuPathTM 0.2.2. Software. The lymphocytes were counted in spleen, thymus, and bursa of Fabricius. The congestion percentage was determined in the spleen and thymus. The interfollicular space in the bursa of Fabricius was also determined.
Immunological and hematological parameters
The antibody titer against NDV was determined by performing the hemagglutination and hemagglutination inhibition tests as previously described [37]. The phagocytic activity of the macrophages present in the blood of the infected birds was determined by a carbon clearance assay, as previously described [38]. 1 mL of Pelican® Black Indian No. 4001 was injected into the wing vein of the birds. 0.2 mL of blood was collected at 0, 3, and 15 minutes intervals and added to 4 mL of 0.1% sodium citrate solution in a 15 mL falcon tube. Centrifuged at 6,000×g for 4 minutes. 50 µL of supernatant was transferred to a 96 well plate, and the optical density value was determined at 650 nm. The lymphoproliferative response against avian tuberculin was determined as previously described [39] by injecting 0.1 mL avian tuberculin into the interdigital space of the right claw of the bird and 0.1 mL normal saline into the interdigital space of the left claw and comparing their immune responses. The hematological parameters (complete blood count) were determined as previously described [40].
Statistical analysis
The statistical analysis of the collected data was performed using the complete randomized design through the analysis of variance technique and Tukey’s test was performed for the comparison of the group mean values using SAS® University edition online software SAS 15.1. p values < 0.01 and < 0.05 were considered significantly different.
RESULTS
Hematological parameters at day 7 and 11 post-infection
A complete blood count analysis of the blood samples infected from S. gallinarum was performed to find the effects of ZnO and CuO nanoparticles. The antibacterial effect of different levels of the ZnO and CuO nanoparticles and florfenicol on the S. gallinarum induced infection in the broilers in terms of the hematological parameters is presented in Table 1.
Table 1.
Antibacterial effect of various levels of mixed zinc oxide (ZnO) and copper oxide (CuO) nanoparticles, and florfenicol against Salmonella gallinarum induced infection in broiler in terms of hematological parameters at day 7 and 11 post-infection
| Hematological Parameters |
Treatments |
p- value |
| Control negative |
Control positive |
Florfenicol (mg/L) |
Nanoparticle levels of ZnO and CuO (mg/kg/d) |
| T1 (50) |
T2 (25 + 10) |
T3 (37.5 + 15) |
T4 (50 + 20) |
| S1 |
| TEC (×106 /µL) |
5.57 ± 0.70a |
4.04 ± 0.30a |
3.81 ± 0.47a |
4.5 ± 0.47a |
4.98 ± 1.60a |
4.6 ± 0.33a |
0.098 |
| TLC (×103 /µL) |
4.47 ± 0.51a |
11.9 ± 0.98b |
7.03 ± 0.55c |
10.03 ± 0.81bd |
9.16 ± 0.72cd |
8.15 ± 1.55cd |
0.000 |
| Hematocrit level (%) |
32.25 ± 1.84a |
22.6 ± 3.54b |
27.5 ± 3.67ab |
24.0 ± 2.44b |
26.0 ± 4.35ab |
22.7 ± 2.52b |
0.003 |
| Hemoglobin concentration (µg/dL) |
10.4 ± 0.60a |
8.76 ± 0.46b |
10.3 ± 3.67a |
8.4 ± 0.84b |
8.1 ± 0.58b |
7.8 ± 0.20b |
0.000 |
| Lymphocytes (%) |
55.02 ± 5.80a |
31.34 ± 3.18b |
49.67 ± 2.33a |
48.7 ± 3.25a |
51.74 ± 5.31a |
50.9 ± 1.55a |
0.000 |
| Heterophils (%) |
29.13 ± 1.60a |
42.22 ± 4.86b |
34.19 ± 2.29a |
34.21 ± 1.54a |
33.04 ± 4.57a |
34.98 ± 2.76ab |
0.006 |
| Monocytes (%) |
5.55 ± 0.58a |
15.29 ± 1.55b |
8.97 ± 1.07ac |
13.68 ± 4.86bc |
11.73 ± 3.39bc |
11.29 ± 0.52ac |
0.001 |
| Eosinophils (%) |
0.53 ± 0.04a |
1.29 ± 0.25a |
1.65 ± 0.18ab |
2.58 ± 0.80b |
2.74 ± 0.96b |
1.79 ± 0.29ab |
0.000 |
| Basophils (%) |
1.16 ± 0.25a |
0.51 ± 0.13a |
0.68 ± 0.02a |
0.81 ± 0.50a |
0.74 ± 0.78a |
1.01 ± 0.62a |
0.636 |
| Heterophil lymphocyte ratio |
0.53 ± 0.09a |
1.35 ± 0.27b |
0.68 ± 0.02a |
0.7 ± 0.04a |
0.64 ± 0.14a |
0.68 ± 0.07a |
0.000 |
| MCH (pg) |
18.72 ± 1.71a |
21.7± 1.42ab |
27.22 ± 2.91b |
18.78 ± 3.47a |
17.01 ± 4.91a |
16.9 ± 1.48a |
0.004 |
| MCV (fL) |
58.33 ± 9.78a |
55.79 ± 5.13a |
73.04 ± 16.65a |
53.58 ± 8.76a |
53.61 ± 9.74a |
49.27 ± 8.09a |
0.540 |
| MCHC (g/dL) |
32.34 ± 3.53a |
39.13 ± 6.90a |
37.89 ± 6.61a |
35.01 ± 1.91a |
31.43 ± 3.50a |
34.63 ± 3.06a |
0.255 |
| S2 |
| TEC (×106 /µL) |
5.6 ± 0.70a |
4.2 ± 0.25a |
4.44 ± 0.73a |
4.38 ± 1.69a |
3.97 ± 1.98a |
3.99 ± 0.63a |
0.162 |
| TLC (×103 /µL) |
6.09 ± 0.69a |
6.1 ± 0.43a |
5.24 ± 0.21a |
4.94 ± 0.74a |
4.87 ± 1.43a |
6.0 ± 0.46a |
0.068 |
| Hematocrit level (%) |
30.75 ± 4.04a |
29.0 ± 3.60a |
25.0 ± 6.63a |
32.8 ± 2.32a |
29.0 ± 2.44a |
28.0 ± 1.73a |
0.379 |
| Hemoglobin concentration (µg/dL) |
10.2 ± 0.34a |
8.93 ± 0.46a |
10.25 ± 0.46a |
10.24 ± 0.77a |
9.2 ± 1.23a |
10.35 ± 0.46a |
0.065 |
| Lymphocytes (%) |
56.04 ± 5.33a |
31.54 ± 3.04b |
49.09 ± 5.49a |
49.0 ± 4.38a |
49.98 ± 0.74a |
48.96 ± 4.69a |
0.000 |
| Heterophils (%) |
25.37 ± 2.67a |
49.09 ± 5.49b |
33.11 ± 1.73c |
34.48 ± 2.00c |
34.96 ± 2.01c |
37.06 ± 3.55bc |
0.000 |
| Monocytes (%) |
8.1 ± 1.65a |
13.51 ± 2.59b |
8.78 ± 2.33ab |
12.06 ± 2.70ab |
11.19 ± 2.11ab |
10.74 ± 1.32ab |
0.030 |
| Eosinophils (%) |
0.79 ± 0.13ab |
1.89 ± 0.38b |
2.03 ± 0.09ac |
3.21 ± 0.68c |
2.42 ± 0.74ac |
1.87 ± 0.69ab |
0.000 |
| Basophils (%) |
1.26 ± 0.40a |
0.92 ± 0.12a |
0.71 ± 0.06a |
1.23 ± 0.31a |
1.44 ± 0.94a |
1.35 ± 0.49a |
0.224 |
| Heterophil lymphocyte ratio |
0.68 ± 0.07a |
0.45 ± 0.09b |
0.67 ± 0.05ac |
0.71 ± 0.09ac |
0.69 ± 0.03ac |
0.76 ± 0.13c |
0.000 |
| MCH (pg) |
18.37 ± 2.83a |
21.29 ± 1.83a |
23.42 ± 4.59a |
24.56 ± 7.82a |
23.93 ± 6.30a |
26.19 ± 3.77a |
0.198 |
| MCV (fL) |
55.69 ± 13.46a |
69.05 ± 8.87a |
58.36 ± 27.39a |
78.66 ± 24.27a |
75.97 ± 23.06a |
71.16 ± 13.67a |
0.406 |
| MCHC (g/dL) |
33.4 ± 3.58a |
31.0 ± 2.44a |
42.32 ± 10.38a |
31.26 ± 2.73a |
31.75 ± 3.90a |
37.02 ± 2.58a |
0.114 |
Download Excel Table
On day 7 post-infection, the influence of the various levels of ZnO and CuO nanoparticles (T2, T3, T4) on the total erythrocyte count, basophils, mean corpuscular volume, and mean corpuscular hemoglobin concentration showed no significant differences with that of the control negative and group T1. The total leukocyte count of T3 and T4 was not significantly different from group T1, however, significantly different (p< 0.01) from control positive, while that of T2 was significantly different (p< 0.01) as compared to T1. The hematocrit level of group T1 and groups T2, T3, and T4 was not significantly different. The lymphocyte, heterophil, monocyte, and eosinophil percentages, and heterophils to lymphocyte (H/L) ratio of groups T2, T3, and T4 were not significantly different from that of group T1, however, lymphocyte percentage of T2, T3, and T4 was found significantly different (p< 0.01) as compared to that of control positive. At day 11 post-infection, the influence of the various levels of ZnO and CuO nanoparticles (T2, T3, and T4) on the total erythrocyte count, total leukocyte count, hematocrit level, hemoglobin concentration, basophils, mean corpuscular hemoglobin, mean corpuscular volume, and mean corpuscular hemoglobin concentration was not significantly different from that of groups; control negative, control positive, and T1. The heterophil and eosinophil percentage of groups; T2 and T3 were significantly different (p < 0.01) as compared to that of control positive. The heterophil, monocyte, and eosinophil percentages of groups T2, T3, and T4 were not significantly different from that of group T1. The lymphocyte percentage and H/L ratio of groups; T2, T3 and T4 was found significantly different (p < 0.01) to that of control positive and not significantly different to that of groups; control negative and T1.
Immunological parameters
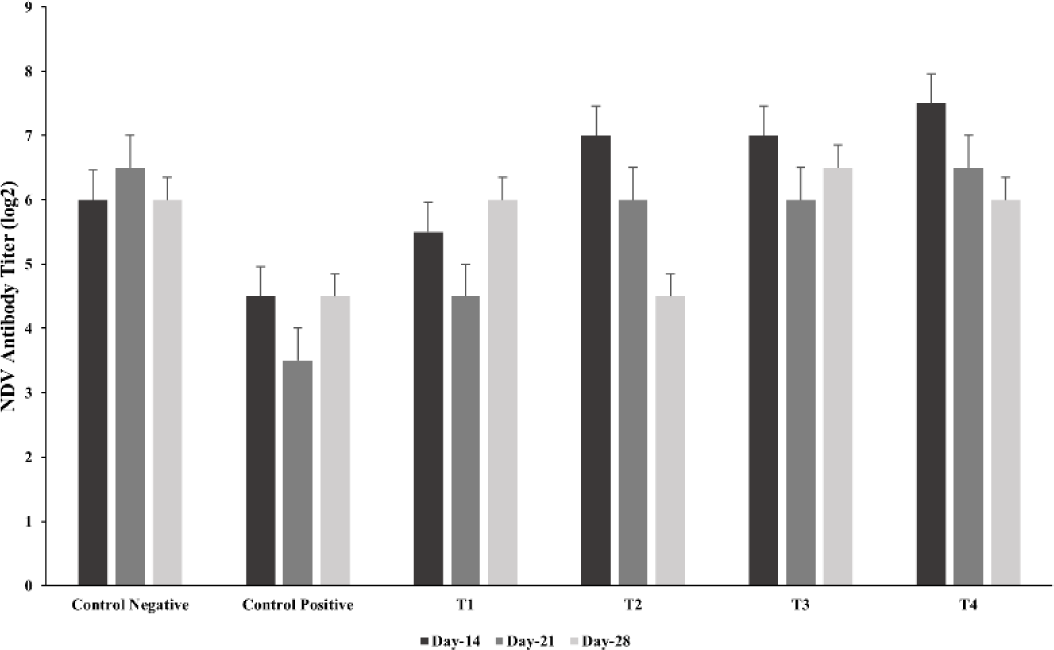
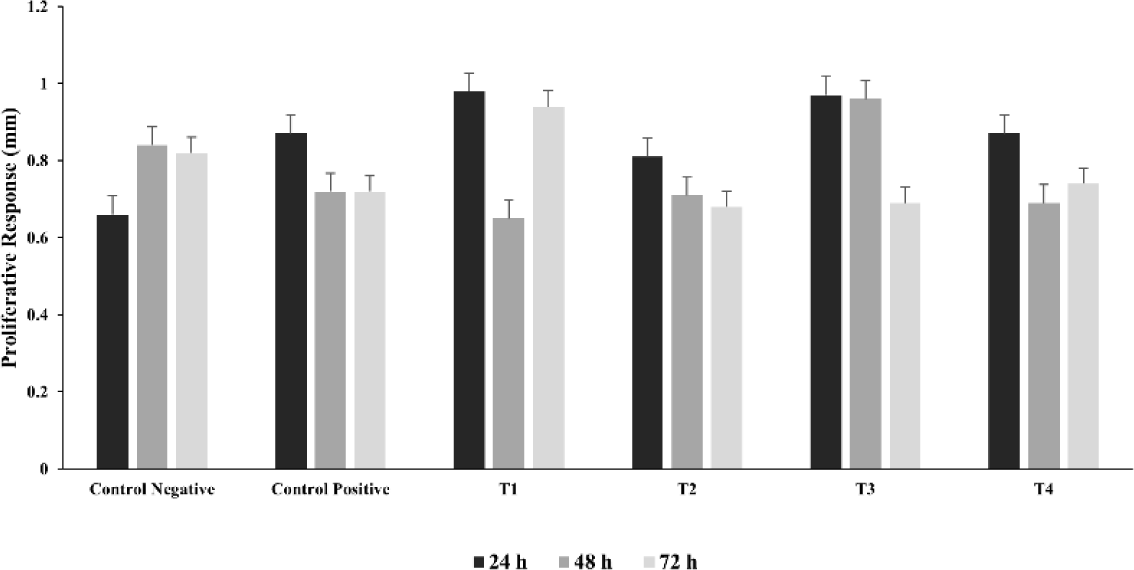
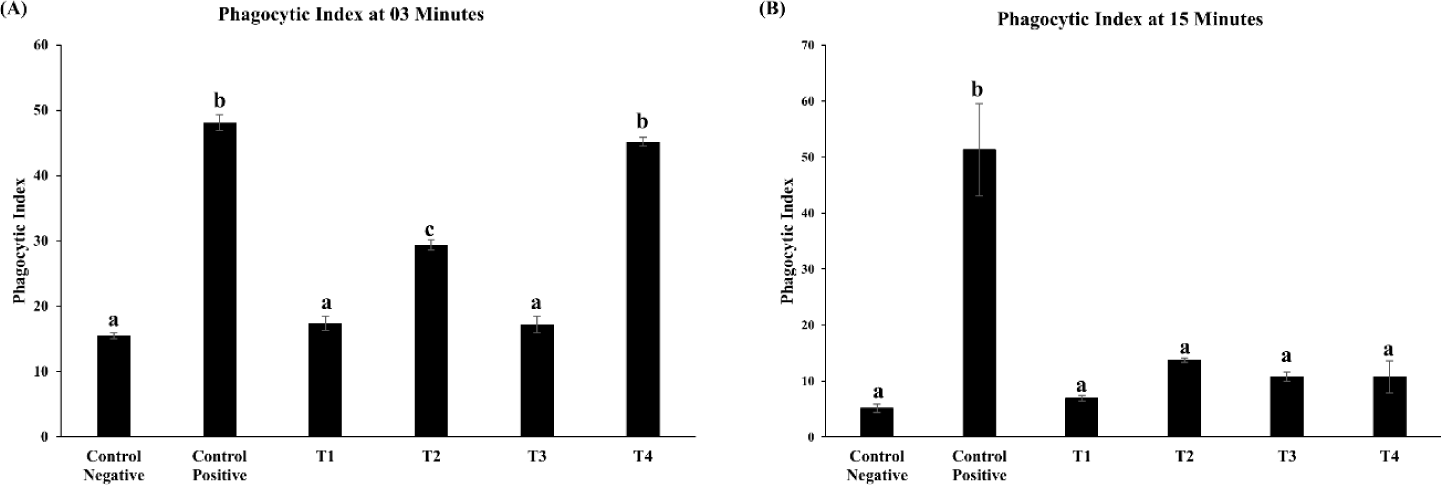
The influence of ZnO and CuO nanoparticles on the immune parameters of the S. gallinarum infected birds was evaluated in terms of antibody titer against NDV, lymphoproliferative response of lymphocytes, and phagocytic power of the macrophages. The log antibody titer against NDV (on days 14, 21, and 28) and lymphoproliferative response of lymphocytes against avian tuberculin (at 24, 48 and 72 hours post injection of avian tuberculin) was not significantly different in the treatment groups (T1, T2, T3, and T4) including the control negative and control positive as described in Figs. 2 and 3 respectively. After 3 minutes, the phagocytic index was significantly (p< 0.01) decreased in the lower and medium treatment groups T2 and T3 as compared to that of control positive group and not significantly different to that of T1 as depicted in Fig. 4A. However, after 15 minutes, the phagocytic index in all groups treated with ZnO and CuO nanoparticles was significantly (p< 0.01) decreased as compared to that of control positive group and not different to that of T1 as depicted in Fig. 4B.
Fig. 2.
New castle’s disease (NDV) titer of Salmonella gallinarum infected birds treated with florfenicol and zinc oxide (ZnO), and copper oxide (CuO) nanoparticles.
Groups: Control negative (no infection, no treatment); control positive (S. gallinarum infection, no treatment); T1 (S. gallinarum infection and florfenicol treatment at dose rate 50 mg/L in drinking water); T2 (S. gallinarum infection and ZnO + CuO nanoparticles treatment at dose rate 25 + 10 mg/kg/d); T3 (S. gallinarum infection and ZnO + CuO nanoparticles treatment at dose rate 37.5 + 15 mg/kg/d); T4 (S. gallinarum infection and ZnO + CuO nanoparticles treatment at dose rate 37.5 + 15 mg/kg/d). Mean ± SD, n = 3 each group.
Download Original Figure
Fig. 3.
Lymphoproliferative response (skin thickness in mm) to injection avian tuberculin in Salmonella gallinarum infected birds treated with florfenicol, zinc oxide (ZnO), and copper oxide (CuO) nanoparticles.
Groups: Control negative (no infection, no treatment); control positive (S. gallinarum infection, no treatment); T1 (S. gallinarum infection and florfenicol treatment at dose rate 50 mg/L in drinking water); T2 (S. gallinarum infection and ZnO + CuO nanoparticles treatment at dose rate 25 + 10 mg/kg/d); T3 (S. gallinarum infection and ZnO + CuO nanoparticles treatment at dose rate 37.5 + 15 mg/kg/d); T4 (S. gallinarum infection and ZnO + CuO nanoparticles treatment at dose rate 37.5 + 15 mg/kg/d). Mean ± SD, n = 3 each group.
Download Original Figure
Fig. 4.
Phagocytic activity of lymphocytes via carbon clearance assay ofSalmonella gallinarum infected birds treated with florfenicol, zinc oxide (ZnO), and copper oxide (CuO) nanoparticles.
(A) Phagocytic index at 3 minutes. (B) Phagocytic index at 15 minutes. Groups: Control negative (no infection, no treatment); COntrol positive (S. gallinarum infection, No Treatment); T1 (S. gallinarum infection and florfenicol treatment at dose rate 50 mg/L in drinking water); T2 (S. gallinarum infection and ZnO + CuO nanoparticles treatment at dose rate 25 + 10 mg/kg/d); T3 (S. gallinarum infection and ZnO + CuO nanoparticles treatment at dose rate 37.5 + 15 mg/kg/d); T4 (S. gallinarum infection and ZnO + CuO nanoparticles treatment at dose rate 37.5 + 15 mg/kg/d). Mean ± SD, n = 3 each group. Values with different letters (a–c) indicate a significantly difference (p < 0.01) and (p < 0.05) phagocytic index.
Download Original Figure
Pathological parameters
Gross pathology of spleen, thymus, bursa of Fabricius
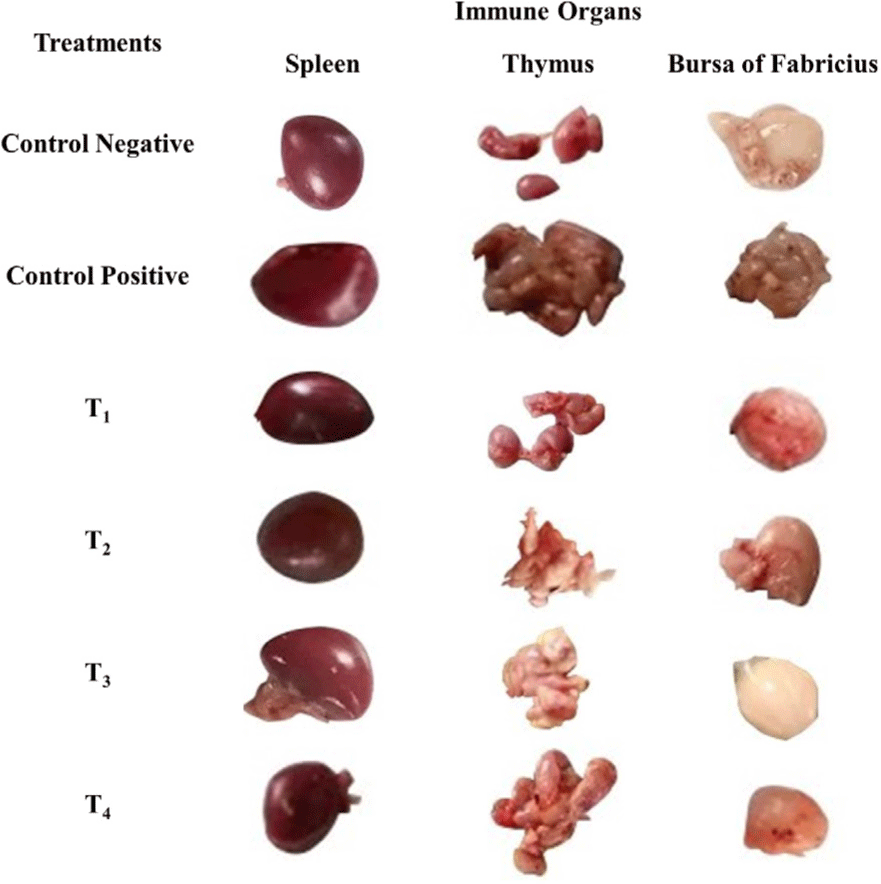
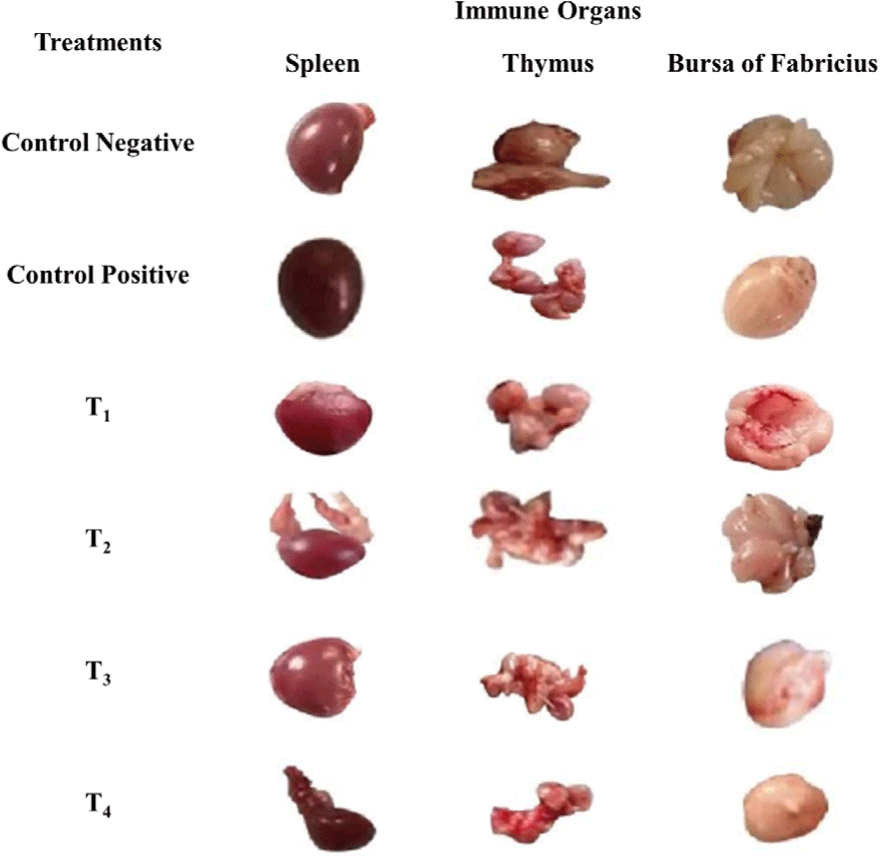
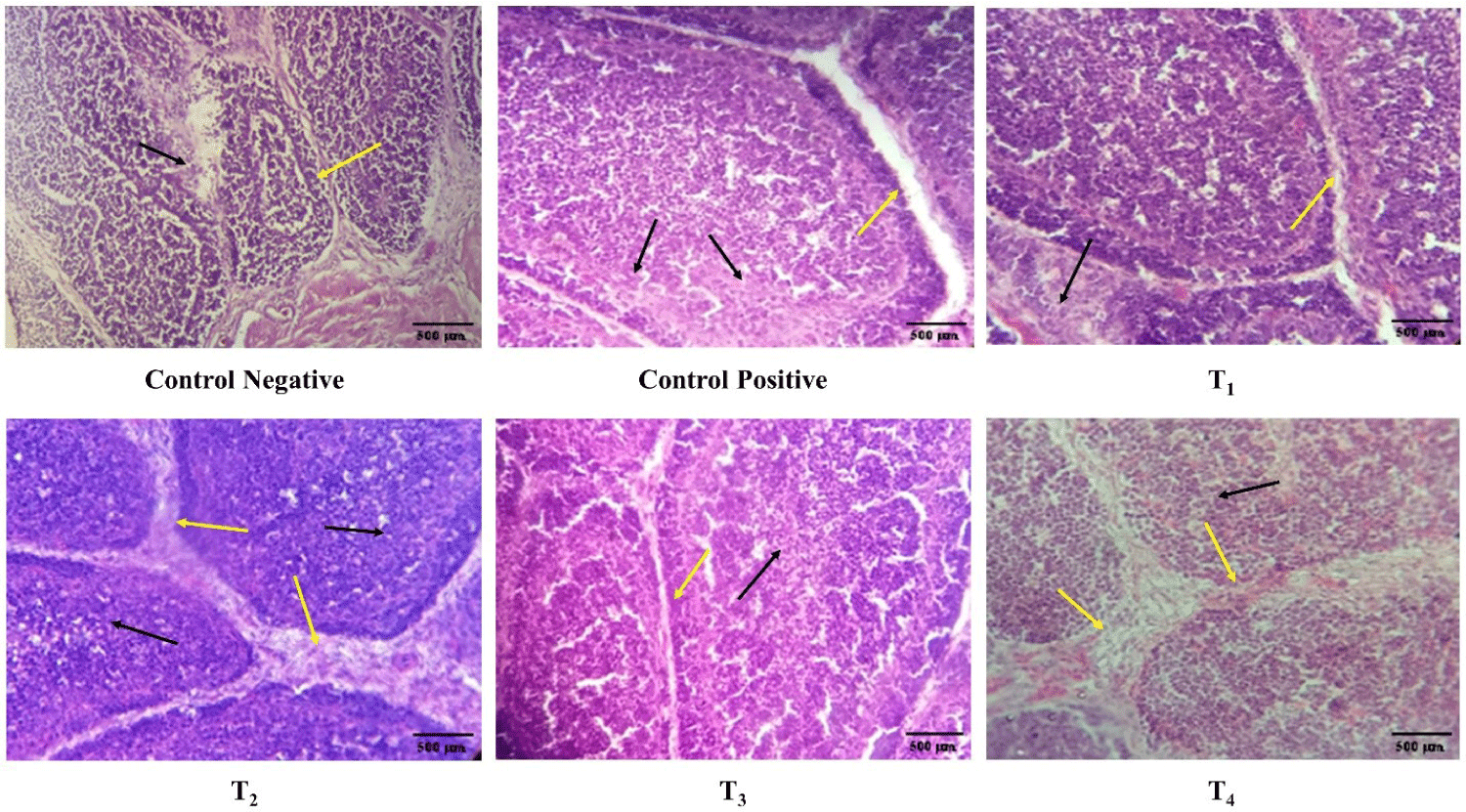
The scoring of congestion of spleen, thymus, and bursa of Fabricius are shown in Table 2, Figs. 5 and 6. At day 7 and 11 post-infection, the congestion in the spleen and thymus of the control positive was high in comparison to the control negative and treatment groups (T1, T2, T3, and T4) as shown in Figs. 5 and 6.
Table 2.
Antibacterial effect of various levels of mixed zinc oxide (ZnO) and copper oxide (CuO) nanoparticles, and florfenicol on Salmonella gallinarum induced infection in broiler in terms of scoring of gross pathological lesions (congestion) of thymus, spleen, and bursa of Fabricius at day 7 and 11 post-infection
| Organ |
Treatments |
| Control negative |
Control positive |
Florfenicol (mg/L) |
Nanoparticle levels of ZnO and CuO (mg/kg/d) |
| T1 (50) |
T2 (25 + 10) |
T3 (37.5 + 15) |
T4 (50 + 20) |
| S1 |
| Spleen |
− |
+++ |
++ |
+ |
++ |
++ |
| Thymus |
− |
+++ |
++ |
++ |
++ |
+ |
| Bursa of Fabricius |
− |
++ |
− |
+ |
++ |
+ |
| S2 |
| Spleen |
− |
++ |
− |
+ |
− |
− |
| Thymus |
− |
++ |
− |
+ |
− |
− |
| Bursa of Fabricius |
− |
++ |
− |
− |
− |
− |
Download Excel Table
Fig. 5.
Antibacterial effect of various levels of mixed zinc oxide (ZnO) and copper oxide (CuO) nanoparticles, and florfenicol on Salmonella gallinarum induced infection in broiler in terms of gross pathology of spleen, thymus, and bursa of Fabricius at day 7 post-infection.
Groups: Control negative (no infection, no treatment); control positive (S. gallinarum infection, no treatment); T1 (S. gallinarum infection and florfenicol treatment at dose rate 50 mg/L in drinking water); T2 (S. gallinarum infection and ZnO + CuO nanoparticles treatment at dose rate 25 + 10 mg/kg/d); T3 (S. gallinarum infection and ZnO + CuO nanoparticles treatment at dose rate 37.5 + 15 mg/kg/d); T4 (S. gallinarum infection and ZnO + CuO nanoparticles treatment at dose rate 37.5 + 15 mg/kg/d).
Download Original Figure
Fig. 6.
Antibacterial effect of various levels of mixed zinc oxide (ZnO) and copper oxide (CuO) nanoparticles, and florfenicol on Salmonella gallinarum induced infection in broiler in terms of gross pathology of spleen, thymus, and bursa of Fabricius at day 11 post-infection.
Groups: Control negative (no infection, no treatment); control positive (S. gallinarum infection, no treatment); T1 (S. gallinarum infection and florfenicol treatment at dose rate 50 mg/L in drinking water); T2 (S. gallinarum infection and ZnO + CuO nanoparticles treatment at dose rate 25 + 10 mg/kg/d); T3 (S. gallinarum infection and ZnO + CuO nanoparticles treatment at dose rate 37.5 + 15 mg/kg/d); T4 (S. gallinarum infection and ZnO + CuO nanoparticles treatment at dose rate 37.5 + 15 mg/kg/d).
Download Original Figure
Histopathology of spleen, thymus, bursa of Fabricius
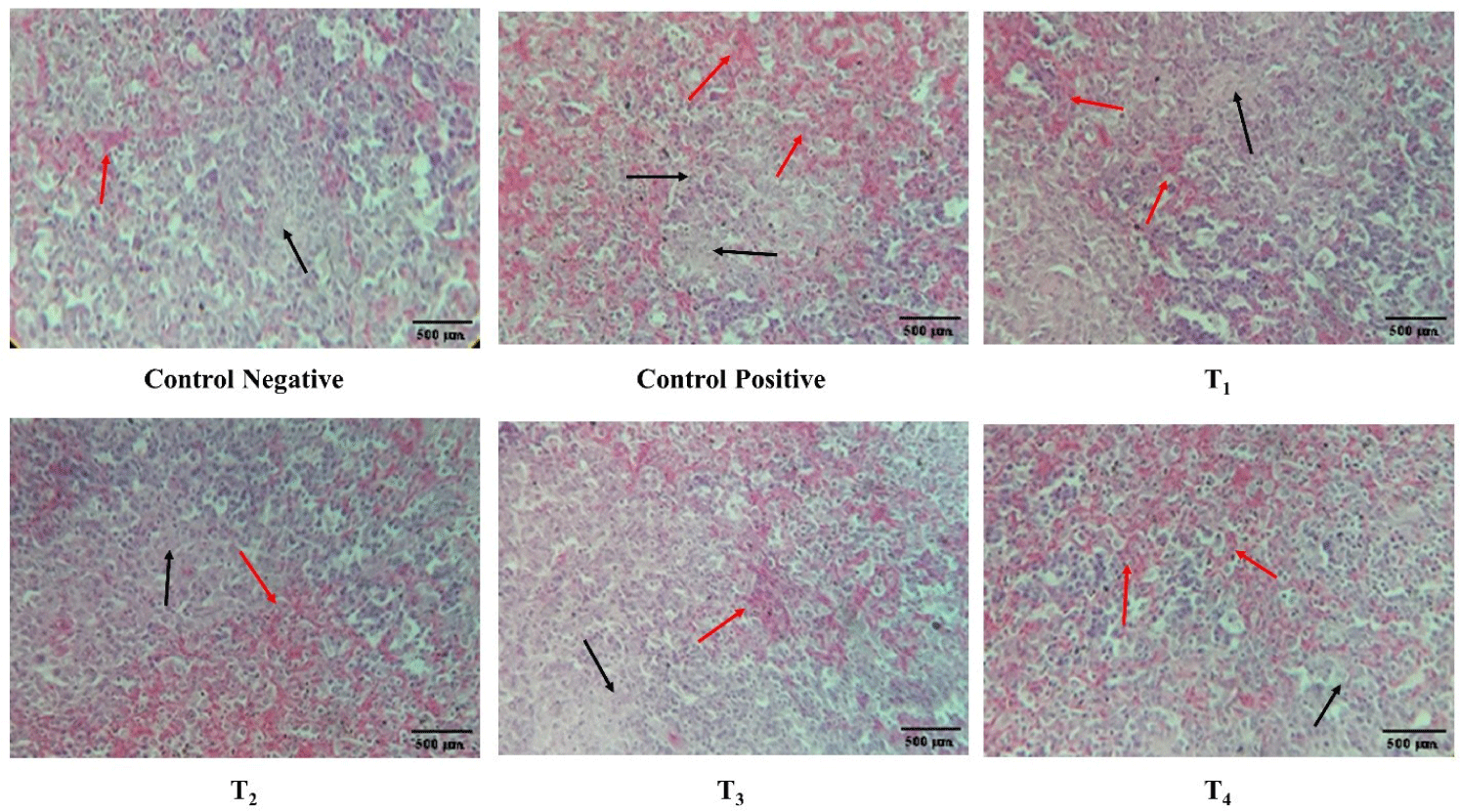
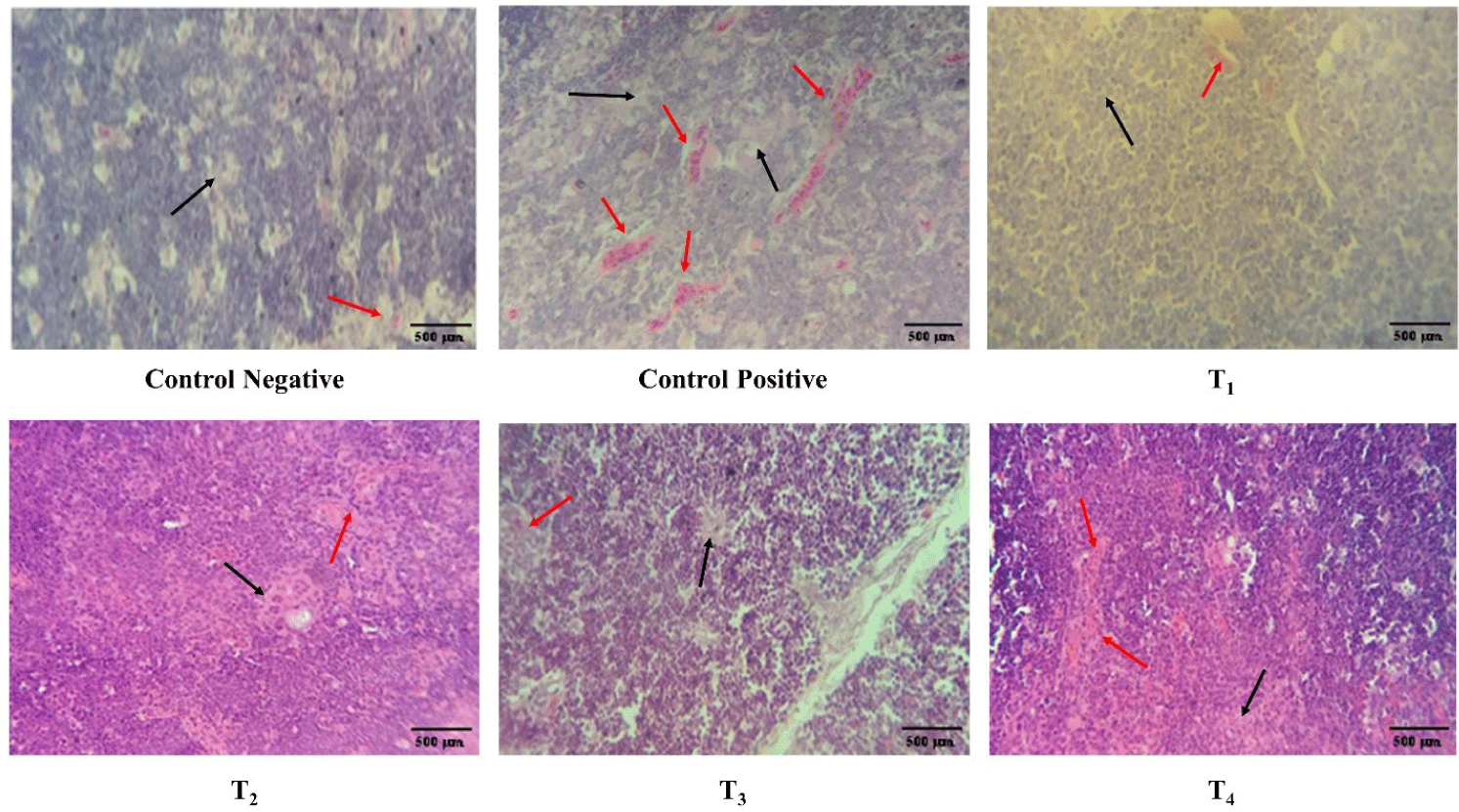
The congestion, lymphocyte count, and interfollicular bursal space in spleen, thymus, and bursa of Fabricius at day 7 and 11 post-infection is described in Table 3 and Figs. 7, 8, and 9, respectively. At day 7 post-infection, the congestion and lymphocytic count in the spleen and thymus of the ZnO and CuO nanoparticles treated groups; T2, T3, and T4 was not significantly different from that of the control negative and T1, however, the congestion and lymphocytic count of spleen in ZnO and CuO nanoparticles treated groups; T2, T3, and T4 were significantly (p< 0.01) different from that of the control positive. ZnO and CuO nanoparticles treated groups; T2, T3, and T4 were significantly different from that of control positive group in terms of congestion (p< 0.01) and lymphocytic count (p< 0.05). The interfollicular space of bursa of Fabricius in ZnO and CuO nanoparticles treated groups was significantly (p< 0.01) different from that of control positive group, however, it was found not different to that T1. At day 11 post-infection, the congestion in the spleen and thymus of ZnO and CuO nanoparticles treated groups (T2, T3, and T4) was not significantly different from that of the control negative and T1, however, was significantly (p< 0.01) different from that of the control positive (Table 3). Congestion and lymphocytic depletion in the spleen and thymus of the control positive was observed while the ZnO and CuO nanoparticles treated groups showed decreased (p< 0.01) congestion of spleen and lymphocytic depletion (Figs. 7 and 8).
Table 3.
Antibacterial effect of various levels of mixed zinc oxide (ZnO) and copper oxide (CuO) nanoparticles, and florfenicol on Salmonella gallinarum induced infection in broiler in terms of quantitative histopathology of thymus, spleen, and bursa of Fabricius at day 7 and day 11 post-infection
| Organ |
Parameters |
Treatment |
p- value |
| Control negative |
Control positive |
Florfenicol (mg/L) |
Nanoparticle levels of ZnO and CuO (mg/kg/d) |
| T1 (50) |
T2 (25 + 10) |
T3 (37.5 + 15) |
T4 (50 + 20) |
| S1 |
| Spleen |
Congestion (%) |
4.33 ± 7.50a |
38.02 ± 7.52b |
20.62 ± 1.14c |
13.38 ± 0.73ac |
10.24 ± 0.48ac |
13.5 ± 2.74ac |
0.000 |
| Lymphocytes |
1,179.67 ± 165.32a |
631.33 ± 178.55b |
983.33 ± 40.69a |
957.67 ± 40.00a |
1,121.67 ± 137.90a |
1,085 ± 58.59a |
0.001 |
| Thymus |
Congestion (%) |
6.00 ± 2.00a |
41.42 ± 6.30b |
19.12 ± 5.80c |
20.65 ± 5.07c |
14.6 ± 1.80ac |
22.34 ± 1.51c |
0.000 |
| Lymphocytes |
1,221.67 ± 241.61a |
703.33 ± 85.13b |
1,055 ± 279.77ab |
915 ± 112.80ab |
1,011.33 ± 26.00ab |
1,020 ± 118.29ab |
0.051 |
| Bursa of Fabricius |
Interfollicular space |
1.06 ± 0.12a |
3.98 ± 0.30b |
2.66 ± 1.07ab |
3.98 ± 0.75b |
2.7 ± 0.81ab |
3.1 ± 0.58b |
0.002 |
| Lymphocytes |
1,103 ± 197.40a |
662.66 ± 86.10b |
858 ± 115.90ab |
968 ± 53.20ab |
994 ± 55.60ab |
996.67 ± 148.90a |
0.012 |
| S2 |
| Spleen |
Congestion (%) |
3.66 ± 2.08a |
41.99 ± 9.37b |
15.38 ± 5.56a |
15.51 ± 3.19a |
13.07 ± 1.23a |
12.5 ± 1.23a |
0.000 |
| Lymphocytes |
1,331.67 ± 334.33a |
862.66 ± 92.52a |
1,130.33 ± 390.37a |
959.33 ± 50.00a |
1,039.67 ± 187.70a |
962 ± 43.55a |
0.276 |
| Thymus |
Congestion (%) |
3.66 ± 2.08a |
41.99 ± 9.37b |
15.38 ± 5.56a |
15.51 ± 3.19a |
13.07 ± 1.23a |
12.5 ± 1.23a |
0.000 |
| Lymphocytes |
1,192.33 ± 335.90a |
700 ± 92.70a |
1,109.67 ± 401.50a |
973 ± 106.60a |
1,057 ± 139.20a |
1,036.33 ± 192.30a |
0.268 |
| Bursa of Fabricius |
Interfollicular space |
0.94 ± 0.10a |
4.09 ± 0.30b |
2.38 ± 1.20ab |
2.68 ± 1.10ab |
2.17 ± 0.10ab |
2.34 ± 0.10ab |
0.003 |
| Lymphocytes |
1,133 ± 352.20a |
747 ± 78.00a |
927.66 ± 64.40a |
630.67 ± 393.90a |
960 ± 175.00a |
1,102.33 ± 283.80a |
0.200 |
Download Excel Table
Fig. 7.
Antibacterial effect of various levels of mixed zinc oxide (ZnO) and copper oxide (ZnO) nanoparticles, and florfenicol on Salmonella gallinarum induced infection in broiler in terms of histopathology of spleen.
Red arrow indicates congestion black arrow indicates lymphocytic depletion. Groups: Control negative (no infection, no treatment); control positive (S. gallinarum infection, no treatment); T1 (S. gallinarum infection and florfenicol treatment at dose rate 50 mg/L in drinking water); T2 (S. gallinarum infection and ZnO + CuO nanoparticles treatment at dose rate 25 + 10 mg/kg/d); T3 (S. gallinarum infection and ZnO + CuO nanoparticles treatment at dose rate 37.5 + 15 mg/kg/d); T4 (S. gallinarum infection and ZnO + CuO nanoparticles treatment at dose rate 37.5 + 15 mg/kg/d); H & E stain, Magnification 10x.
Download Original Figure
Fig. 8.
Antibacterial effect of various levels of mixed zinc oxide (ZnO) and copper oxide (CuO) nanoparticles, and florfenicol onSalmonella gallinarum induced infection in broiler in terms of histopathology of thymus. Red arrow indicates congestion, black arrow indicates lymphocytic depletion.
Groups: Control negative (no infection, no treatment); control positive (S. gallinarum infection, no treatment); T1 (S. gallinarum infection and florfenicol treatment at dose rate 50 mg/L in drinking water); T2 (S. gallinarum infection and ZnO + CuO nanoparticles treatment at dose rate 25 + 10 mg/kg/d); T3 (S. gallinarum infection and ZnO + CuO nanoparticles treatment at dose rate 37.5 + 15 mg/kg/d); T4 (S. gallinarum infection and ZnO + CuO nanoparticles treatment at dose rate 37.5 + 15 mg/kg/d); H & E stain, Magnification 10x.
Download Original Figure
Fig. 9.
Antibacterial effect of various levels of mixed zinc oxide (ZnO) and copper oxide (CuO) nanoparticles, and florfenicol on Salmonella gallinarum induced infection in broiler in terms of histopathology of bursa of Fabricius.
Yellow arrow indicates interfollicular space, black arrow indicates lymphocytic depletion. Groups: Control negative (no infection, no treatment); control positive (S. gallinarum infection, no treatment); T1 (S. gallinarum infection and florfenicol treatment at dose rate 50 mg/L in drinking water); T2 (S. gallinarum infection and ZnO + CuO nanoparticles treatment at dose rate 25 + 10 mg/kg/d); T3 (S. gallinarum infection and ZnO + CuO nanoparticles treatment at dose rate 37.5 + 15 mg/kg/d); T4 (S. gallinarum infection and ZnO + CuO nanoparticles treatment at dose rate 37.5 + 15 mg/kg/d); H & E stain, Magnification 10x.
Download Original Figure
DISCUSSION
Fowl typhoid, caused by gram-negative bacterium S. gallinarum, poses a significant economic burden on the global poultry industry [28]. Multiple antibiotics such as florfenicol, ciprofloxacin, and enrofloxacin are being used against S. gallinarum at poultry farms however, the irrational use of these antibiotics created AMR in S. gallinarum. Morsy et al. [41] reported that ZnO and CuO nanoparticles have no cytotoxic effects in the broiler chickens at low doses, however, at high dose it can cause cytotoxicity. However, in another study reported that ZnO, CuO, and Ferric oxide nanocomposite can ameliorate the toxic effects of ochratoxins in broilers and can improve the body weight, liver, and kidney functions [42]. To address the issue of AMR, this study aimed to assess the antibacterial activity of ZnO and CuO nanoparticles against S. gallinarum infection in broiler chicken.
The findings of this study highlight the antibacterial potential of nanoparticles as a significant alternative treatment approach for combating S. gallinarum infection in broiler chickens. In response to S. gallinarum infection at day 19 with 108 CFU/mL, the clinical signs like huddling, anorexia, depression, pasty yellow diarrhea, and postmortem lesions including bronzed colored liver, splenomegaly, necrotic foci on the visceral organs (liver and heart), and mortality greater than 60% in the birds were observed. These findings are consistent with previous studies [43–45] where they reported 50% mortality, rough feathers, yellow-green diarrhea, and sunken eyes. The bursa of Fabricius is very important and unique immune organ responsible for B lymphocytes production and humoral immunity in birds [46]. Therefore, the bursa of Fabricius was observed and based on evaluation it was noted that after nanoparticle treatment no congestion in the bursa of Fabricius was observed. The treatment of nanoparticles can increase the relative weight of bursa of Fabricius [47]. The recovery of the bursa of Fabricius from congestion was attributed to the antibacterial activity of the combination of nanoparticles [44].
The total erythrocytic count, total leukocyte count, hemoglobin concentration, and hematocrit level were found similar in all groups which was also reported in previous studies [48,49]. The intraperitoneal administered infection of S. gallinarum causes a significant decrease in hematocrit and hemoglobin concentration as compared to per oral infection [46]. The hematocrit and hemoglobin was decreased in the S. gallinarum infected birds as compared to the control negative which was in agreement with another study [50]. An arithmetic decrease in erythrocyte count and substantial decrease in hemoglobin concentration and hematocrit of the S. gallinarum infected birds was observed. The decreased hemoglobin and hematocrit level caused anemia in the birds [51]. The anemia in the control positive might be attributed to the increased ability of the reticuloendothelial system to take up modified erythrocytes [52]. The increase in the erythrocyte count in the nanoparticles treated groups may be attributed to the efficacy of CuO nanoparticles, because copper plays a vital role in iron metabolism for hemoglobin synthesis [53] and erythrocyte production [54]. After inducing S. gallinarum infection, an increased total leukocyte count was observed in the infected groups which is in line with a previous study [50] because they play a key role in the defense mechanism of the host and active removal of the bacteria from circulation [55]. The increase in leukocyte count indicates the severity of infection while CuO nanoparticles reduces the leukocyte count in the blood [56]. After the treatment with ZnO and CuO nanoparticles, total leukocyte counts decreased. However, the total leukocyte count in the group T3 (day 7 post-infection) was found comparable with that of T1 group treated with florfenicol antibiotic, whereas the leukocyte number was not different with that of the control negative at the second sampling at day 11 post-infection, which indicates that nanoparticles have similar efficacy as the antibiotics.
In contrast, Fathi et al. [48] obtained contrasting findings where they observed that nanoparticles had no impact on the leukocytic count. This discrepancy might be attributed to the induced S. gallinarum infection in our study as bacterial infections cause an increase in leukocytic count [57]. An increase in the leukocyte count of infected birds and the substantial decrease due to nanoparticle treatment is due to the adequate efficacy of the nanoparticles as they contribute to reducing the leukocytic count [58]. Ahmed et al. [6] also reported that the CuO nanoparticles decreased the leukocyte count in birds infected with E. coli and elaborates on the efficacy of ZnO and CuO nanoparticles against S. gallinarum infection in broilers. A significant increase in heterophil percentage was observed in the control positive as compared to the control negative. The nanoparticles treatment can decrease the heterophils percentage. Heterophilia in the control positive could be an indication of acute inflammatory degenerative changes in the internal organs [43]. The stress can also be linked with the impaired immunity of the birds. The infection of S. gallinarum can cause the increased level of corticosterone [59]. The heterophils percentage can be increased with increased level of corticosterone in the blood [60]. However, the treatment groups T1, T2, and T3 showed a significantly decreased heterophil percentage as compared to the control positive, while the heterophil percentage in the groups (T2, T3, and T4) treated with nanoparticles was found comparable to group T1. However, group T3 showed a lower heterophil percentage as compared to T2 and T4 indicating that the group T3 may have the minimum inflammatory degenerative changes which endorse the efficacy of treatment with ZnO and CuO nanoparticles at the levels of 37.5 + 15 mg/kg/d. At day 7 post-infection, the decreased H/L ratio in the nanoparticle-treated groups might be attributed to the efficacy of nanoparticles in the alleviation of stress due to the S. gallinarum infection. The leukocytosis and heterophilia in the control positive could be due to the inflammatory response to the S. gallinarum induced tissue damage. The decreased leukocyte count and heterophil percentage in the nanoparticle-treated groups indicates the improved health status of the birds and antibacterial activity of ZnO and CuO nanoparticles which cause a decrease in bacterial load and inflammatory degeneration in the infected birds.
The CuO nanoparticles can inhibit the growth of NDV [61]. The nonsignificant difference between all the treatments in NDV antibody titer at day 14, 21, and 27 in the current study was endorsed by previous in vivo studies [62]. On the other hand, a previous study reported that the humoral immune response was increased when using ZnO nanoparticles [63]. The nonsignificant titers against NDV in the present study could be due to the induced S. gallinarum infection. The NDV antibody titer was highest in group T2 receiving a low dose of nanoparticles and lowest in group T4 receiving a high dose of nanoparticles which is in line with the previous study reported by Morsy et al. [41]. The low NDV antibody titer in group T4 may be attributed to the oxidative stress induced by the high level of nanoparticles [64].
The macrophage phagocytic activity of the nanoparticle-treated groups T2, T3, and T4 were enhanced. Macrophages are involved in the initiation of cellular and humoral immune responses by activating the B and T lymphocytes [38]. Copper plays an important role in the production of arachidonic acid and prostaglandin which enhances the production of macrophages [65]. The minimum light absorption percentage in T3 indicates the increased phagocytic activity of macrophages which may be attributed to the efficacy of the nanoparticles dose level in T3. The decreased light absorption in the nanoparticle-treated groups demonstrates the increased phagocytic activity of the macrophages indicating improved immune status of the nanoparticle-treated groups.
The histopathological examination of spleen, thymus, and bursa of Fabricius in our study indicated lymphocytic depletion and congestion in the S. gallinarum infected birds which also endorsed by a previous in vivo study [66]. The histological sections of spleen, thymus and bursa of Fabricius showed lymphocytic depletion and congestion which is in line with the previous studies [67–69]. The lymphocytic depletion in the S. gallinarum infected groups may be attributed to immunosuppression in the presence of the S. gallinarum induced infection [50]. In our study, the lymphocyte count in spleen, thymus and bursa of Fabricius was increased while the congestion was decreased after treatment with ZnO and CuO nanoparticles in the S. gallinarum induced infected birds which indicates the efficacy of the nanoparticle treatment.
In conclusion, ZnO and CuO nanoparticles at the dose level of 37.5 + 15 mg/kg/d and 50 + 20 mg/kg/d, respectively, showed optimum therapeutic activity against S. gallinarum infection in broilers. As the two dose levels show equal therapeutic results against S. gallinarum, the lower dose 37.5 + 15 mg/kg/d recommended.
Acknowledgements
The author(s) are thankful to Dr. Yasir Javed and Dr. Sohail Sajid (University of Agriculture Faisalabad) for providing chemicals and shed facility for the experimental trials. The author(s) are also thankful to Dr. Manshaad Basheer (Big Bird, Pakistan) for providing experimental chicks for this study.
Availability of data and material
Ethics approval and consent to participate
REFERENCES
Ghafoor A, Badar H, Hussain M, Tariq N. An empirical estimation of the factors affecting demand and supply of poultry meat. Pak Vet J. 2010; 30:172-4

Cisman MM, Ahmed ZA, Mohamoud HA, Awale AT, Nour HSH. Scope specification of coccidiosis in the poultry on researchers. Int J Avian Wildlife Biol. 2020; 5:32-7

Mottet A, Tempio G. Global poultry production: current state and future outlook and challenges. Worlds Poult Sci J. 2017; 73:245-56
Mack S, Hoffmann D, Otte J. The contribution of poultry to rural development. Worlds Poult Sci J. 2005; 61:7-14
Alikhan NF, Moreno LZ, Castellanos LR, Chattaway MA, McLauchlin J, Lodge M, et al. Dynamics of Salmonella enterica and antimicrobial resistance in the Brazilian poultry industry and global impacts on public health. PLOS Genet. 2022; 18e1010174
Ahmed MH, Javed MT, Bahadur SUK, Tariq A, Tahir MH, Tariq ME, et al. Antibacterial effects of copper oxide nanoparticles against E. coli induced infection in broilers. Appl Nanosci. 2022; 12:2031-44
Abbas G, Khan SH, Hassan M, Mahmood S, Naz S, Gilani SS. Incidence of poultry diseases in different seasons in Khushab district, Pakistan. J Adv Vet Anim Res. 2015; 2:141-5
Berhanu G, Fulasa A. Pullorum disease and fowl typhoid in poultry: a review. Br J Poult Sci. 2020; 9:48-56
Ahmed MS, Sarker A, Rahman MM. Prevalence of infectious diseases of broiler chickens in Gazipur district. Bangladesh J Vet Med. 2009; 7:326-31
Rashid MH, Xue C, Islam MR, Islam MT, Cao Y. A longitudinal study on the incidence of mortality of infectious diseases of commercial layer birds in Bangladesh. Prev Vet Med. 2013; 109:354-8
Jawale CV, Chaudhari AA, Lee JH. Generation of a safety enhanced Salmonella Gallinarum ghost using antibiotic resistance free plasmid and its potential as an effective inactivated vaccine candidate against fowl typhoid. Vaccine. 2014; 32:1093-9
Mbuko IJ, Raji MA, Ameh J, Saidu L, Musa WI, Abdul PA. Prevalence and seasonality of fowl typhoid disease in Zaria-Kaduna State, Nigeria. J Bacteriol Res. 2009; 1:001-5
Poole T, Sheffield C. Use and misuse of antimicrobial drugs in poultry and livestock: mechanisms of antimicrobial resistance. Pak Vet J. 2013; 33:266-71
Raza MA, Javed MT, Fiaz M, Shakeel M, Haq MSU, Kanwal A, et al. Antibacterial effect of zinc oxide and copper oxide nanoparticles as substitute of antibiotics against fowl typhoid in broilers. Pak J Zool. 2022; :1-9
Oloso NO, Adeyemo IA, van Heerden H, Fasanmi OG, Fasina FO. Antimicrobial drug administration and antimicrobial resistance of salmonella isolates originating from the broiler production value chain in Nigeria. Antibiotics. 2019; 8:75
de Mesquita Souza Saraiva M, Lim K, do Monte DFM, Givisiez PEN, Alves LBR, de Freitas Neto OC, et al. Antimicrobial resistance in the globalized food chain: a One Health perspective applied to the poultry industry. Braz J Microbiol. 2022; 53:465-86
Mouttotou N, Ahmad S, Kamran Z, Koutoulis KC. Prevalence, risks and antibiotic resistance of Salmonella in poultry production chain.In In: Mares M, editor.editor Current topics in Salmonella and Salmonellosis. Rijeka: IntechOpen. 2017; p p. 215-34
Threlfall EJ. Antimicrobial drug resistance in Salmonella: problems and perspectives in food- and water-borne infections. FEMS Microbiol Rev. 2002; 26:141-8
White DG, Zhao S, Simjee S, Wagner DD, McDermott PF. Antimicrobial resistance of foodborne pathogens. Microbes Infect. 2002; 4:405-12
Naik AB, Selukar NB. Role of nanotechnology in medicine. Everyman’s Sci. 2009; 44:151-3
Azam A, Ahmed AS, Oves M, Khan MS, Habib SS, Memic A. Antimicrobial activity of metal oxide nanoparticles against Gram-positive and Gram-negative bacteria: a comparative study. Int J Nanomedicine. 2012; 7:6003-9
Hameed ASH, Karthikeyan C, Ahamed AP, Thajuddin N, Alharbi NS, Alharbi SA, et al. In vitro antibacterial activity of ZnO and Nd doped ZnO nanoparticles against ESBL producing Escherichia coli and Klebsiella pneumoniae. Sci Rep. 2016; 6:24312
Khan MI, Fatima N, Shakil M, Tahir MB, Riaz KN, Rafique M, et al. Investigation of in-vitro antibacterial and seed germination properties of green synthesized pure and nickel doped ZnO nanoparticles. Phys B Condens Matter. 2021; 601:412563
Mohamed AA, Abu-Elghait M, Ahmed NE, Salem SS. Eco-friendly mycogenic synthesis of ZnO and CuO nanoparticles for in vitro antibacterial, antibiofilm, and antifungal applications. Biol Trace Elem Res. 2021; 199:2788-99
Kim T, Kim M, Lee J, Moturi J, Ha S, Tajudeen H, et al. Supplementation of nano-zinc in lower doses as an alternative to pharmacological doses of ZnO in weanling pigs. J Anim Sci Technol. 2022; 64:70-83
Slavin YN, Asnis J, Häfeli UO, Bach H. Metal nanoparticles: understanding the mechanisms behind antibacterial activity. J Nanobiotechnol. 2017; 15:65
Blecher K, Nasir A, Friedman A. The growing role of nanotechnology in combating infectious disease. Virulence. 2011; 2:395-401
Xie Y, He Y, Irwin PL, Jin T, Shi X. Antibacterial activity and mechanism of action of zinc oxide nanoparticles against Campylobacter jejuni. Appl Environ Microbiol. 2011; 77:2325-31
Dadi R, Azouani R, Traore M, Mielcarek C, Kanaev A. Antibacterial activity of ZnO and CuO nanoparticles against gram positive and gram negative strains. Mater Sci Eng C. 2019; 104:109968
Lee J, Hosseindoust A, Kim M, Kim K, Kim T, Moturi J, et al. Effects of hot-melt extruded nano-copper on the Cu bioavailability and growth of broiler chickens. J Anim Sci Technol. 2021; 63:295-304
Kim B, Jeong JY, Park SH, Jung H, Kim M. Effects of dietary copper sources and levels on growth performance, copper digestibility, fecal and serum mineral characteristics in growing pigs. J Anim Sci Technol. 2022; 64:885-96
Kim M, Cho JH, Seong PN, Jung H, Jeong JY, Kim S, et al. Fecal microbiome shifts by different forms of copper supplementations in growing pigs. J Anim Sci Technol. 2021; 63:1386-96
Phiwdang K, Suphankij S, Mekprasart W, Pecharapa W. Synthesis of CuO nanoparticles by precipitation method using different precursors. Energy Procedia. 2013; 34:740-5
Manyasree D, Kiran acid P, Venkata RK. Characterization and antibacterial activity of ZnO nanoparticles synthesized by co precipitation method. Int J Appl Pharm. 2018; 10:224-8
Zarnab S, Javed MT, Gul AHST, Mahmood MS. The chicken in-house environment can be improved by the use of nanotechnology. Park Vet J. 2022; 42:526-32
Bancroft JD, Stevens A. Theory and practice of histological techniques. 4th ed New York, London and Madrid: Churchill Livingstone. 1996
Swayne DE, Glisson JR, Jackwood MW, Pearson JE, Reed WM. A laboratory manual for the isolation and identification of avian
pathogens. 4th ed Kennett Square, PA: American Association of Avian Pathologists. 1998
Sarker N, Tsudzuki M, Nishibori M, Yasue H, Yamamoto Y. Cell-mediated and humoral immunity and phagocytic ability in chicken lines divergently selected for serum immunoglobulin M and G levels. Poult Sci. 2000; 79:1705-9
Corrier DE. Comparison of phytohemagglutinin-induced cutaneous hypersensitivity reactions in the interdigital skin of broiler and layer chicks. Avian Dis. 1990; 34:369-73
Benjamin MM. Outline of veterinary clinical pathology. 3rd ed Ames, IA: Iowa State University Press. 1978
Morsy EA, Hussien AM, Ibrahim MA, Farroh KY, Hassanen EI. Cytotoxicity and genotoxicity of copper oxide nanoparticles in chickens. Biol Trace Elem Res. 2021; 199:4731-45
Al Shap NF, El-Sherbeny EME, El Masry DMA. The efficacy of metal nanocomposite (Fe3O4/CuO/ZnO) to ameliorate the toxic effects of ochratoxin in broilers. BMC Vet Res. 2022; 18:312
Shah SN, Kamil SA, Darzi MM, Mir MS, Bhat SA. Haematological and some biochemical changes in experimental fowl typhoid infection in broiler chickens. Comp Clin Pathol. 2013; 22:83-91
Kumari D, Mishra SK, Lather D. Pathomicrobial studies on Salmonella Gallinarum infection in broiler chickens. Vet World. 2013; 6:725-9
Chiroma MA, Adamu S, Gadzama JJ, Esievo KAN, Abdulsalam H, Balami AG, et al. Some plasma biochemical changes in layers experimentally infected with Salmonella gallinarum. Afr J Cell Pathol. 2017; 9:66-72
Chen C, Li J, Zhang H, Xie Y, Xiong L, Liu H, et al. Effects of a probiotic on the growth performance, intestinal flora, and immune function of chicks infected with Salmonella pullorum. Poult Sci. 2020; 99:5316-23
Al-Saeedi MKI, Dakhil HH, Al-Khafaji FRA. Effect of adding silver nanoparticles with drinking water on some lymphatic organs and microflora in the intestinal for broiler chickens (ROSS 308). IOP Conf Ser Earth Environ Sci. 2021; 722:012004
Fathi M. Effects of zinc oxide nanoparticles supplementation on mortality due to ascites and performance growth in broiler chickens. Iran J Appl Anim Sci. 2016; 6:389-94
Rezaei A, Farzinpour A, Vaziry A, Jalili A. Effects of silver nanoparticles on hematological parameters and hepatorenal functions in laying japanese Quails. Biol Trace Elem Res. 2018; 185:475-85
Deshmukh S, Asrani RK, Ledoux DR, Rottinghaus GE, Bermudez AJ, Gupta VK. Pathologic changes in extrahepatic organs and agglutinin response to Salmonella Gallinarum infection in Japanese quail fed Fusarium verticillioides culture material containing known levels of fumonisin B1. Avian Dis. 2007; 51:705-12
Prasanna K, Paliwal OP, Kumar R, Tripathi BN. Immunocytochemical detection of Salmonella Gallinarum and Salmonella Pullorum in experimentally infected chickens. Indian J Anim Sci. 2002; 72:113-6
Assoku RKG, Penhale WJ. The anaemia in fowl typhoid: immuno-pathogenesis and associated patterns of erythrocyte destruction. J Comp Pathol. 1978; 88:219-36
Samanta B, Ghosh PR, Biswas A, Das SK. The effects of copper supplementation on the performance and hematological parameters of broiler chickens. Asian-Australas J Anim Sci. 2011; 24:1001-6
Sharma DC, Kochar B, Bhardwaj A, Riyat M, Sharma P. Effect of ingestion of copper bhasm on red cell indices, iron parameters and essential elements in chicks. Indian J Clin Biochem. 2009; 24:245-9
Kokosharov T. Changes in the white blood cells and specific phagocytosis in chicken with experimental acute fowl typhoid. Vet Arhiv. 1998; 68:33-8
Khabbazi M, Harsij M, Akbar Hedayati SA, Gholipoor H, Gerami MH, Farsani HG. Effect of CuO nanoparticles on some hematological indices of rainbow trout Oncorhynchus mykiss and their potential toxicity. Nanomed J. 2015; 2:67-73
Lavoignet CE, Le Borgne P, Chabrier S, Bidoire J, Slimani H, Chevrolet-Lavoignet J, et al. White blood cell count and eosinopenia as valuable tools for the diagnosis of bacterial infections in the ED. Eur J Clin Microbiol Infect Dis. 2019; 38:1523-32
Mroczek-Sosnowska N, Batorska M, Lukasiewicz M, Wnuk A, Sawosz E, Jaworski S, et al. Effect of nanoparticles of copper and copper sulfate administered in ovo on hematological and biochemical blood markers of broiler chickens. Ann Wars Univ Life Sci SGGW Anim Sci. 2013; 52:141-9
Sharma P, Pande VV, Moyle TS, McWhorter AR, Chousalkar KK. Correlating bacterial shedding with fecal corticosterone levels and serological responses from layer hens experimentally infected with Salmonella Typhimurium. Vet Res. 2017; 48:5
Sirirat N, Lu JJ, Hung ATY, Chen SY, Lien TF. Effects different levels of nanoparticles chromium picolinate supplementation on growth performance, mineral retention, and immune responses in broiler chickens. J Agric Sci. 2012; 4:48-58
Yugandhar P, Vasavi T, Jayavardhana Rao Y, Uma Maheswari Devi P, Narasimha G, Savithramma N. Cost effective, green synthesis of copper oxide nanoparticles using fruit extract of Syzygium alternifolium (Wt.) Walp., characterization and evaluation of antiviral activity. J Clust Sci. 2018; 29:743-55
Swart E, Dvorak J, Hernádi S, Goodall T, Kille P, Spurgeon D, et al. The effects of in vivo exposure to copper oxide nanoparticles on the gut microbiome, host immunity, and susceptibility to a bacterial infection in earthworms. Nanomaterials. 2020; 10:1337
Khajeh Bami M, Afsharmanesh M, Salarmoini M, Tavakoli H. Effect of zinc oxide nanoparticles and Bacillus coagulans as probiotic on growth, histomorphology of intestine, and immune parameters in broiler chickens. Comp Clin Pathol. 2018; 27:399-406
Najafi-Hajivar S, Zakeri-Milani P, Mohammadi H, Niazi M, Soleymani-Goloujeh M, Baradaran B, et al. Overview on experimental models of interactions between nanoparticles and the immune system. Biomed Pharmacother. 2016; 83:1365-78
Ricciotti E, FitzGerald GA. Prostaglandins and inflammation. Arterioscler Thromb Vasc Biol. 2011; 31:986-1000
Akter T, Nooruzzaman M, Belal SMSH, Ahammed M, Uddin AJ, Parvin R, et al. Fowl typhoid live lyophilized vaccine applied at 3-month intervals protected layer chickens from Salmonella gallinarum infection and prevented cloacal shedding. J Adv Vet Anim Res. 2022; 9:301-9
Hosen J, Rahman M, Alam J, Das Z, Khan M, Haider M. Pathology of fowl typhoid and molecular detection of its pathogen. Ann Bangladesh Agric. 2019; 23:49-60
Alam J, Chakma T, Islam MS, Islam MT, Khan MAHNA, Islam MT, et al. Pathology of fowl paratyphoid and molecular detection of its pathogen in layer chickens. Ann Bangladesh Agric. 2020; 24:47-57
Gupta S, Jindal N, Khokhar RS, Asrani RK, Ledoux DR, Rottinghaus GE. Individual and combined effects of ochratoxin A and Salmonella enterica serovar Gallinarum infection on pathological changes in broiler chickens. Avian Pathol. 2008; 37:265-72