













INTRODUCTION
in vitro maturation (IVM) is essential for better understanding of process of oocyte development and maturation in various species, including pigs [1]. IVM induces meiotic maturation from prophase I to metaphase II (MII) in vitro. Oocytes generated by IVM are used to produce high-quality embryos upon in vitro fertilization (IVF) and somatic cell nuclear transfer (SCNT). Modulation of follicle size [2,3], medium composition [4,5], culture temperature [6,7], antioxidants [8,9] and cumulus features [10] can enhance the quality of porcine oocytes in IVM. Specifically, supplementation of antioxidant to IVM medium improved embryo quality, reduced early apoptosis levels, increased glutathione (GSH) content and reduced reactive oxygen species (ROS) accumulation, promoting embryonic development [11]. However, further research is needed to improve IVM of porcine oocytes.
Porcine oocytes, IVF embryos, and parthenotes are matured for 42–44 h and SCNT oocytes are matured for 36–38 h in vitro. During this period, most oocytes proceed to metaphase and then to pro-metaphase if the nucleus has sufficiently matured [12,13]. While in meiotic arrest, the nuclear status and structural morphology of matured oocytes remain unchanged. This amount of time is sufficient for oocyte nuclear maturation but not for full maturation of the oocyte cytoplasm [14]. Many studies have been conducted on different maturation starts to acquire oocyte capacitation, which plays an important role in oocytes reaching MII [15–17]. Lin et al. extended the duration of IVM to 52 h in order to increase the poor-quality oocytes and performed treatment with melatonin to inhibit ROS production, apoptosis, and DNA damage [15]. However, oocyte maturation for an excessive amount of time causes cytoplasmic changes that negatively affect oocyte quality and increase the risk of spontaneous oocyte activation [18,19] and subsequent aberrant cleavage characterized by unequally sized blastomeres [19,20]. It is possible to prevent, delay, or reverse these cellular and molecular abnormalities [21].
Rapamycin has antifungal and immunosuppressant properties [22] and binds to FK506-binding protein 12 to form a complex that prevents the kinase activity and function of mTOR [23]. mTOR activity inhibition by rapamycin affects establishment of the cortical granule-free zone and actin cap, and disrupts alignment of the surrounding spindle and division during oocyte meiotic maturation [24]. Administration of rapamycin during IVM of porcine oocytes dose-dependently enhances cytoplasmic and nuclear maturation by inducing autophagy [25]. Aged porcine oocytes treated with 10 μM rapamycin enhances blastocyst quality by regulating the mitochondrial distribution, autophagy, apoptotic cells, and mTOR signaling [26]. Supplementation of tissue culture medium (TCM)-199 with 0.5 μM rapamycin increases expression of matrix metallopeptidase in the trophoblast and inner cell mass (ICM), while it inhibits apoptosis [27]. Therefore, in our study, we extended the IVM duration to fully mature porcine oocytes and performed rapamycin treatment to reduce apoptosis, ROS production, and oocyte aging. This study investigated the impacts of rapamycin treatment during prolonged IVM on molecular maturation factors and the developmental capacity of porcine oocytes in vitro. Activated mitogen-activated protein kinase (MAPK) and ROS levels were modulated upon extension of the IVM duration with rapamycin treatment. The in vitro development rate, total cell numbers, and level of apoptosis were determined using blastocysts generated from these oocytes.
MATERIALS AND METHODS
Unless otherwise specified, all chemicals and reagents utilized in this study were procured from Merck (St. Louis, MO, USA). The oocytes and embryos were maintained in CO2 incubator at 38.8°C in a humidified atmosphere containing 5% CO2 and 95% air.
Porcine ovaries from pre-pubertal sows were obtained from a provincial slaughterhouse. We used an 18-gauge needle attached to 10mL syringe to aspirate follicles measuring 2-8mm from the follicles, and selected cumulus-oocyte-complexes (COCs). After washing in TCM-199-hydroxyethyl piperazine ethane sulfonicacid (HEPES) supplemented with 0.1% (w/v) bovine serum albumin (BSA), a group of approximately 50 selected COCs were cultured in 500 µL of TCM-199 (M-199, Gibco, Grand Island, NY, USA) containing Earle’s salts, 0.57 mM cysteine, 10% (v/v) porcine follicular fluid, 10 ng/mL epidermal growth (E-9644), 0.5 µg/mL follicle-stimulating hormone (F-2293), and 0.5 µg/mL luteinizing hormone (L-5269). The maturation process was conducted for 44 h, with the COCs placed beneath a layer of mineral oil. Thereafter, MII oocytes were transferred into TCM-199 supplemented with 0 or 10 µM rapamycin (R-8781) and incubated for 8 h (total IVM duration of 52 h). According to our previous studies, the concentration of rapamycin was set at the 10 μM [26].
Subsequent IVM, adherent cumulus cells were eliminated using 0.1% (w/v) hyaluronidase. Collected oocytes were parthenogenetically activated by incubation in 5 µM Ca2+ ionomycin (Merck) for 5 min. Following culture in porcine zygote medium-5 (PZM-5) with 7.5 µg/mL cytochalasin B (Merck) for 4 h, the oocytes were washed with PZM-5 containing 0.4% (w/v) BSA. Following 6 days of incubation in the same medium, the oocytes were washed with Dulbecco’s phosphate-buffered saline (PBS). Finally, oocytes and embryos were fixed in 3.7% (w/v) paraformaldehyde for 20 min at 4°C or they were snap freezing using liquid nitrogen and subsequently stored at −70°C.
Intracellular ROS activity in denuded oocytes were quantified using the 2,7-dichlorofluorescein protocol described previously [26]. Concisely, oocytes (25–30 oocytes per sample, four replicates) were cultured in 100 µM 2’,7’-dichlorodihydrofluorescein-diacetate (DCHF-DA) for 20 min, washed five times with PZM-5, and promptly observed under epifluorescence microscopy (Olympus, Tokyo, Japan) with an ultraviolet filter (450–490 nm and 515–565 nm). A microscope-mounted digital camera (Nikon, Tokyo, Japan) was used to capture grayscale images, and ImageJ software (NIH, Bethesda, MD, USA) was employed to acquire mean grayscale values. Image analysis was conducted with the Adobe Photoshop CS6 software package (version 13, Adobe Systems, San Jose, CA, USA) by quantitating the average pixel intensities in various regions of the raw image. Before statistical analysis, background fluorescence values were calculated by subtracting them from the final values.
Blastocysts on day 6 were washed with PBS (pH 7.4) with 1 mg/mL PBS/ polyvinylpyrrolidone (PVP). After fixing with 3.7% formaldehyde prepared with PBS overnight at 4°C. Following washing with PBS/PVP, the oocytes were subsequently permeabilized by culture in the dark for 1 h with 0.3% Triton X-100 at room temperature. After washing with PBS/PVP, the blastocysts were cultured at 37°C for 1 h in fluorescein-conjugated dUTP and terminal deoxynucleotidyl transferase using the In Situ Cell Death Detection Kit (Roche, Mannheim, Germany). Following counterstaining with Hoechst 33342 in the dark for 30 min at 37°C to label the nuclei, the samples were washed with PBS/PVP and then carefully set on glass slides. The experiment was independently repeated three times.
mRNA extraction from oocytes at the MII stage and in vitro-cultured embryos at day 6 (20 embryos per sample, three replicates) by a Dynabeads mRNA Direct Kit (DynalAsa, Oslo, Norway). SuperScript™ III reverse transcriptase (Invitrogen, Grand Island, NY, USA) and oligo (dT)12–18 primers were used to synthesize first-strand cDNA. Real-time quantitative PCR was conducted on a Step One Plus Real-time PCR system (Applied Biosystems, Warrington, UK) using the primers specified in Table 1. The total reaction volume for the final PCR consisted of 20 μL, including SYBR Green PCR Master Mix (Applied Biosystems). As follows were the amplification conditions: 10 min at 94°C, followed by 39 cycles of denaturation for 30 sec at 94°C, annealing for 30 sec at 55°C, and extension for 55 sec at 72°C, and a final extension for 5 min at 72°C. Relative mRNA expression levels were determined according to the 2−∆∆Ct protocol [28] by normalization to GAPDH.
The experimental assay followed a previously described protocol [24]. For protein extraction, 20 µL of 1× sodium dodecyl sulfate (SDS) sample buffer containing 5 mM Tris-HCl, pH 6.8 at 25°C, 2% (w/v) SDS, 10% (v/v) glycerol, 50 mM dithiothreitol (DTT), and 0.01% (w/v) bromophenol blue or phenol red was added to oocytes (20 oocytes per sample, three replicates), and the samples were boiled for 5 min at 95°C. Subsequently, proteins were separated by electrophoresis on a 5%–12% Tris-SDS-polyacrylamide gel electrophoresis (PAGE) gel for 1.5 h at 80–100 V. Thereafter, proteins were electrophoretically transferred to a nitrocellulose membrane (Hybond-ECL, Amersham, Buckinghamshire, UK) at 300 mA for 2 h using transfer buffer (25 mM Tris, 200 mM glycine, and 20% methanol, pH 8.5). After being blocked with 5% (w/v) skim milk prepared in PBS for 1 h, the membranes were cultured with an anti-Cdc2 p34, anti-p44/42 MAPK, or anti-phospho-p44/42 MAPK antibody (1:500; Cell Signaling Technology, Danvers, MA, USA) in blocking solution (1× Tris-buffered saline containing 0.1% (v/v) Tween® 20 and 5% [w/v] skim milk) for at least 2 h. After washing with TBST (20 mM Tris-HCl, pH 7.5, 250 mM NaCl, and 0.1% [v/v] Tween® 20), the membranes were cultured with anti-rabbit IgG-horseradish peroxidase (1:2000, Cell Signaling Technology) with blocking solution for 1 h. Following washing with TBST, binding of antibody was analyzed with chemiluminescence luminol reagent (Invitrogen).
The Statistical Analysis System (SAS User’s Guide; Statistical Analysis System, Cary, NC, USA) was employed to analyze data using the general linear model procedure. Significant differences were determined using Tukey’s multiple range test. Relative gene expression levels were compared by the Student’s t-test. Differences were regarded as biologically significant at p < 0.05 and p < 0.01.
RESULTS
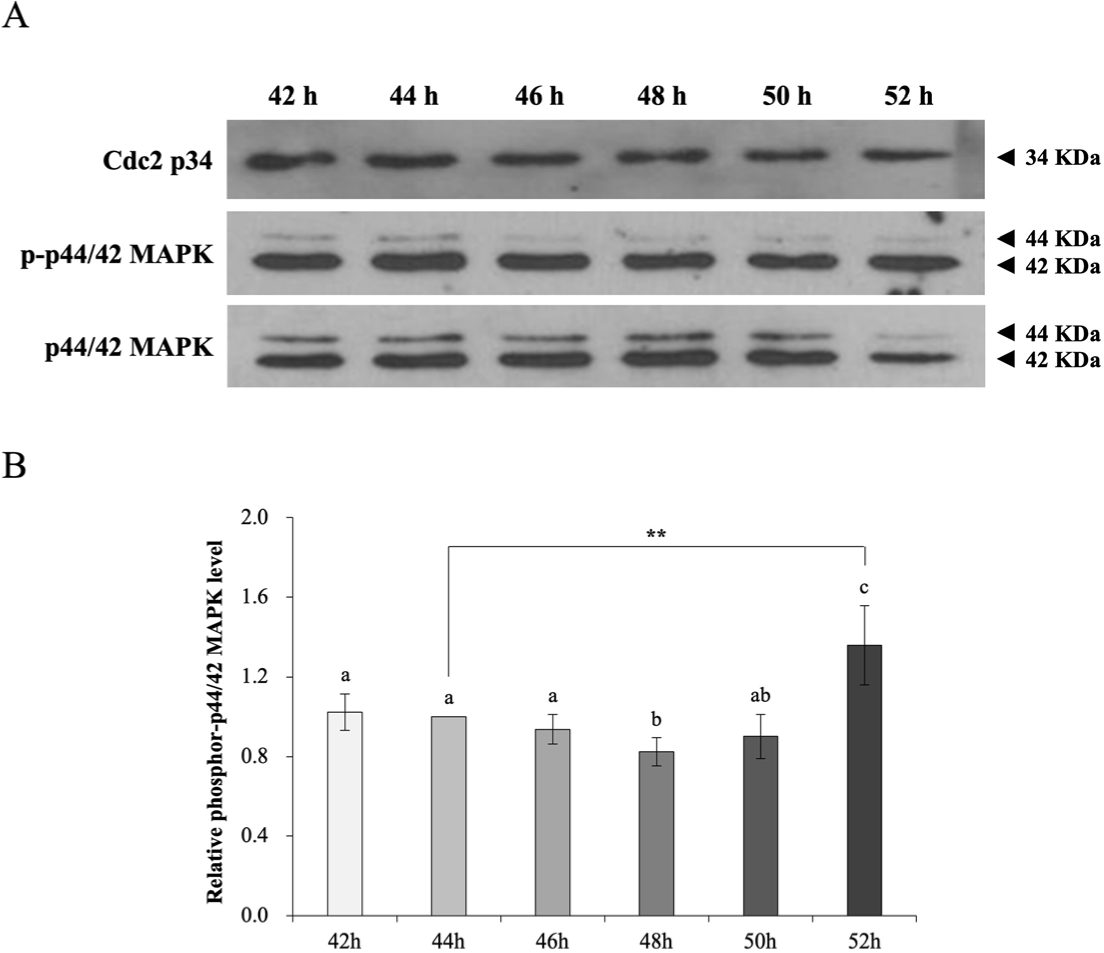
To explore the influence of extension of the IVM duration on molecular maturation of porcine oocytes, we examined expression of maturation/M phase-promoting factor (MPF), activated MAPK, and maternal genes (Figs. 1 and 2). We monitored total and phospho-p44/42 MAPK levels at 2 h intervals after 42–52 h of IVM (Fig. 1A). The phospho-p44/42 MAPK level was normalized against the total MAPK level at 44 h (set to 1). The phospho-p44/42 MAPK level was upregulated in comparison with the total MAPK level at 52 h (42 h, 1.02 ± 0.09; 46 h, 0.94 ± 0.07; 48 h, 0.82 ± 0.07; 50 h, 0.90 ± 0.11; and 52 h, 1.36 ± 0.20; p < 0.05 and p < 0.01; Fig. 1B). The phospho-p44/42 MAPK level was higher at 52 h than at the other time points (Figs. 1A and 1B).
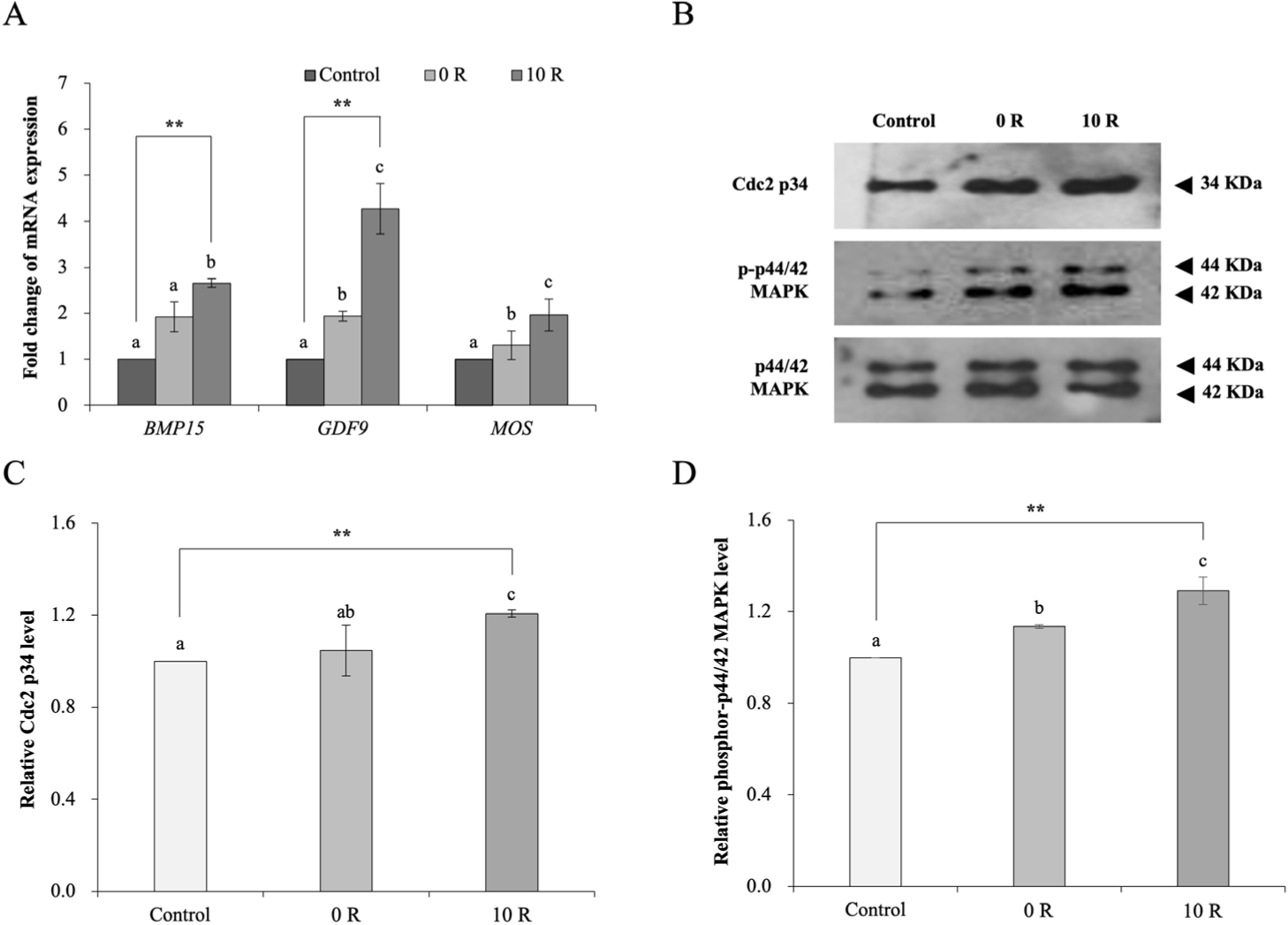
Oocytes were supplemented with and without 10 μM rapamycin and examined after 52 h of IVM (10 R and 0 R, respectively), whereas control oocytes were examined subsequent 44 h of IVM. The mRNA level of the maternal genes BMP15, GDF9, and MOS was analyzed using real-time quantitative PCR (Fig. 2A). The mRNA level of these genes was upregulated in 10 R oocytes compared to control and 0 R oocytes (p < 0.05). There was an increase (p < 0.01) in the levels of BMP15 and GDF9 in 10 R oocytes compared with control oocytes. We monitored the levels of Cdc2 p34 and phospho-p44/42 MAPK (Fig. 2B). Expression of Cdc p34 and phospho-p44/42 MAPK was normalized to the total MAPK in each group (set to 1, Figs. 2C and 2D). The levels of Cdc2 p34 and phospho-p44/42 MAPK were higher in 10 R oocytes (p < 0.01) than in control and 0 R oocytes (0 R, 1.05 ± 0.11 and 1.14 ± 0.01, respectively; and 10 R, 1.21 ± 0.02 and 1.29 ± 0.06, respectively; Figs. 2C and 2D).
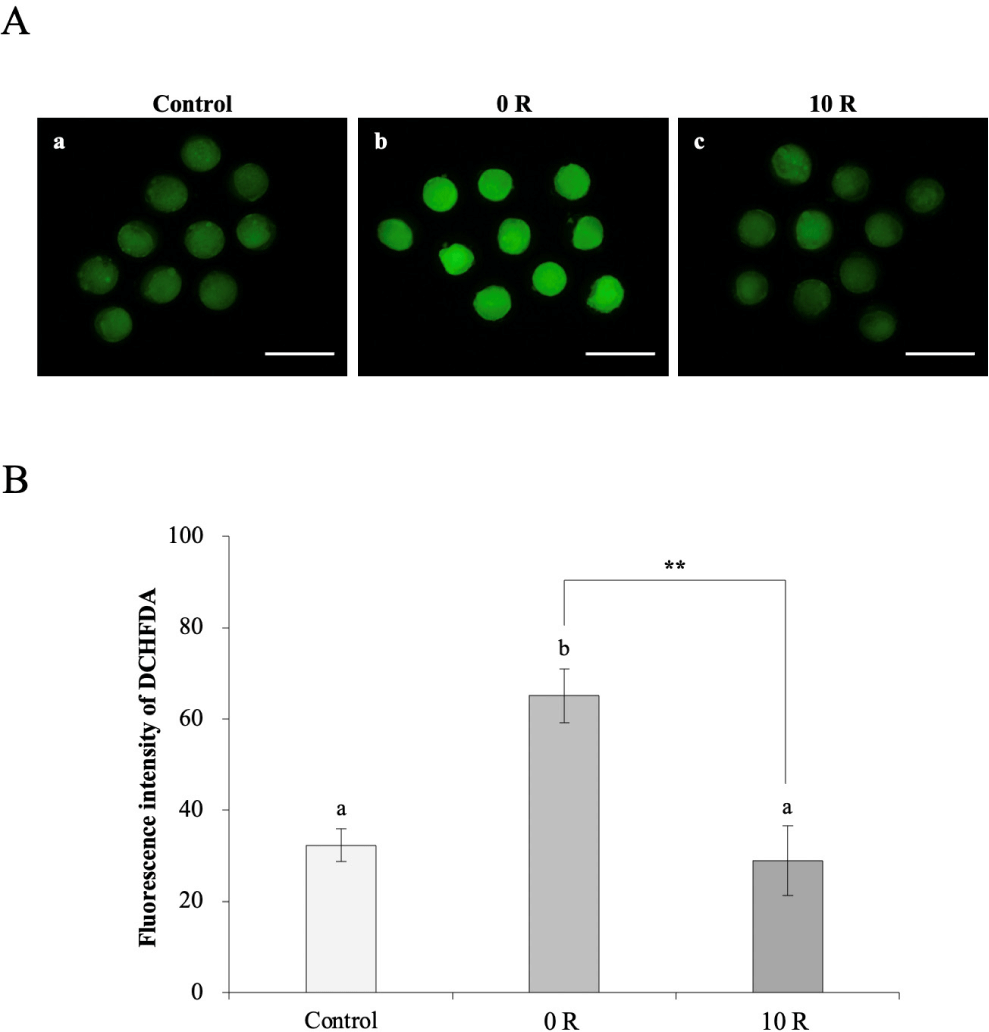
ROS activity was analyzed in control, 0 R, and 10 R groups using dichloro-dihydro-fluorescein diacetate (DCHFDA). ROS activity was lower in 10 R group (p < 0.01) compared to 0 R oocytes and was similar to that in control oocytes (control, 32.3±3.6; 0 R, 65.1±5.9; and 10 R, 28.9±7.6; Figs. 3A and 3B). Treatment with 10 µM rapamycin enhanced the developmental capacity of oocytes aged for 8 h by inhibiting ROS activity.
Control (n = 400) and 0 R (n = 345) and 10 R (n = 310) groups were matured for 44 and 52 h, respectively. The percentage of matured oocytes was no differences among the control, 0 R, and 10 R groups (86.40 ± 1.92%, 90.30 ± 2.03%, and 91.77 ± 2.86%, respectively; Fig. 4B). After IVM, control (n = 345), 0 R (n = 312), and 10 R (n = 284) groups were parthenogenetically activated. The morphology and percentage of embryos reaching the 2–4-cell stage showed no significant difference among the control, 0 R, and 10 R groups (68.73 ± 5.57%, 56.58 ± 5.10%, and 57.61 ± 4.60%, respectively; Figs. 4A and 4B). The percentage of blastocyst at day 6 was highest in the 10 R group (n = 75, 48.36 ± 7.06%) and lowest in the 0 R group (n = 38, 22.81 ± 4.12%; p < 0.05 or p < 0.01; Figs. 4A and 4B).
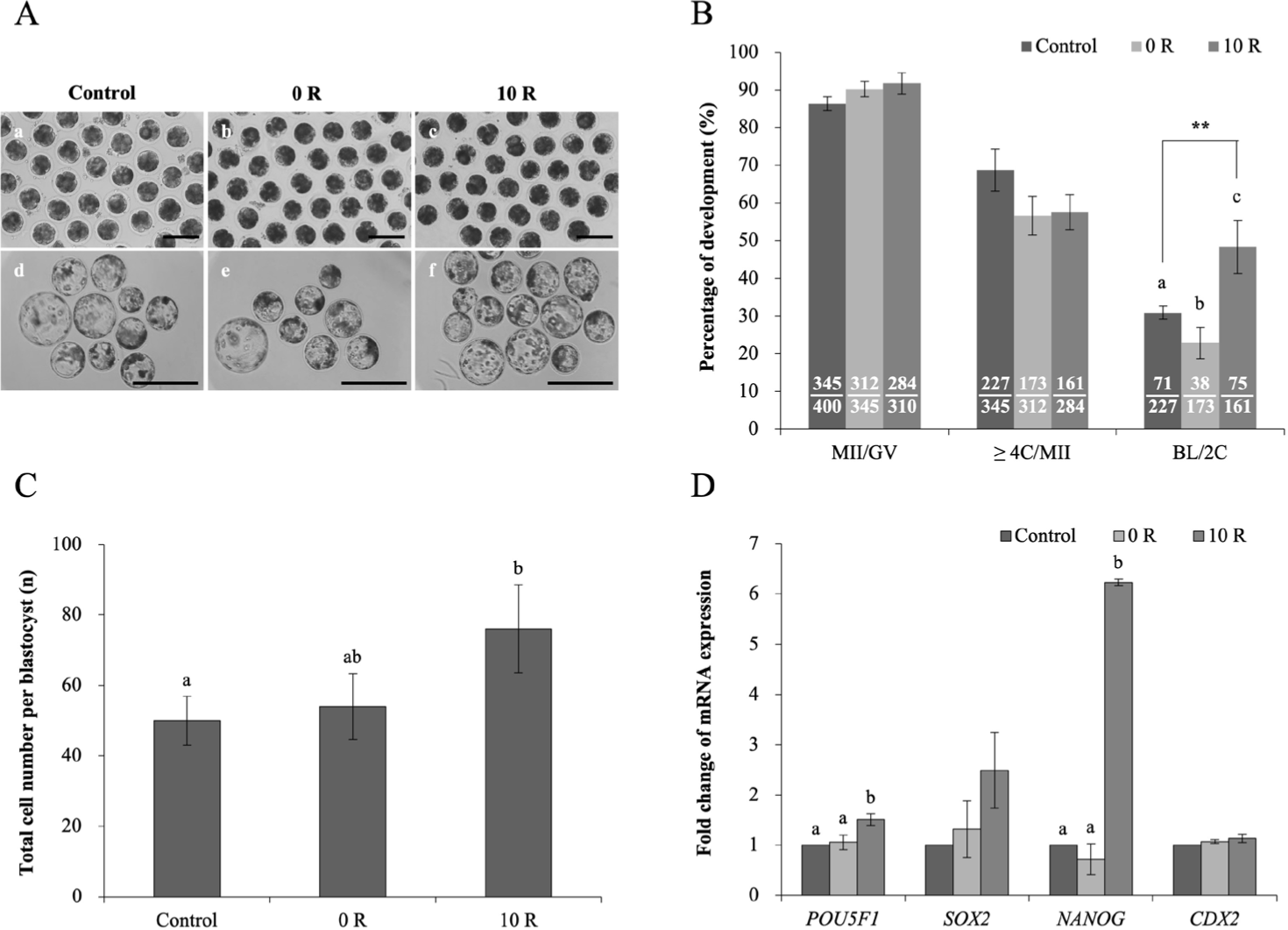
Confirming the influence of rapamycin treatment on blastocyst quality, blastocysts at day 6 in the various groups were stained (Fig. 4C). The cell number per blastocyst at day 6 was highest in the 10 R group (control, 50 ± 6.92; 0 R, 54 ± 9.29; and 10 R, 76 ± 12.49; p < 0.05; Fig. 4C). Reprogramming-related transcription factor genes POU5F1, SOX2, NANOG, and CDX2 were examined for their expression levels by real-time RT-PCR (Fig. 4D). The 10 R group was upregulated (p < 0.05) in POU5F1 and NANOG expression, whereas it did not significant difference among the control and 0 R groups. SOX2 expression tended to show a similar pattern, although there were not significantly differences among the groups. Expression of CDX2 showed no significant difference among the groups.
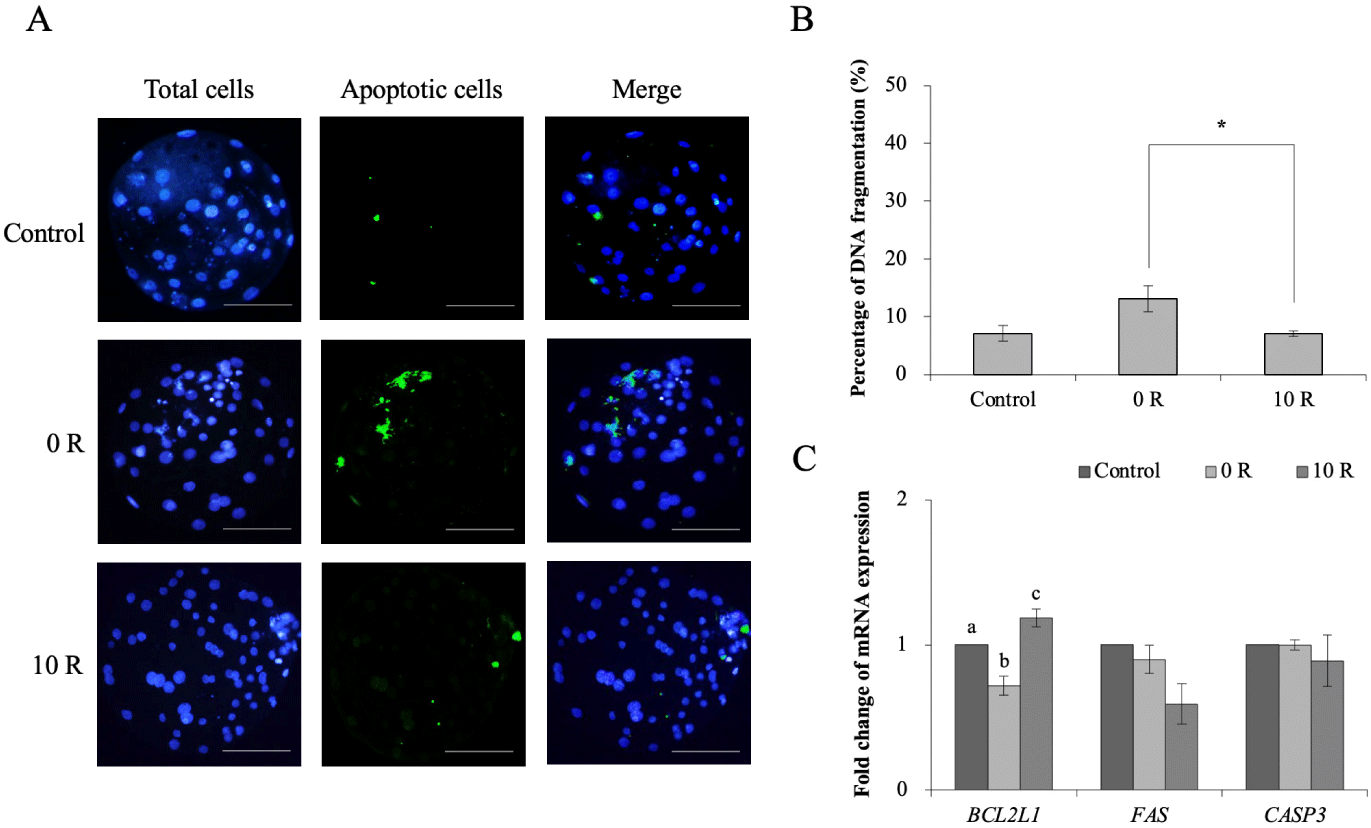
Using the TUNEL assay, individual embryos were assessed for genomic DNA fragmentation (an indicator of apoptosis). The percentage of fragmented DNA in the 10 R group (7.08 ± 0.50%) was significantly downregulated (p < 0.05) compared with the 0 R group (13.06 ± 2.24%) and was similar to the control group (7.07 ± 1.37%, Fig. 5B). The 10 R group showed significantly higher levels of BCL2L1, the anti-apoptotic gene, compared to the 0 R group (p < 0.05), while the control group exhibited the highest expression (p < 0.05) (Fig. 5C). The mRNA level of the FAS and CASP3, pro-apoptotic genes, tended to be downregulated in the 10 R group, although these differences were not significant.
DISCUSSION
Oocyte maturation is a multifaceted process involving both nucleus and cytoplasmic changes. An IVM duration of 42 to 44 h is sufficient for nuclear maturation through a mechanism such as oocyte capacity, but not sufficient for complete maturation of the cytoplasm [14]. However, excessive maturation duration exposes oocytes to increased oxidative stress [29,30]. ROS-induced oocyte damage reduces the ability and quality of subsequent embryonic development [31]. This study investigated the influences of extension of the IVM duration with rapamycin treatment on molecular maturation factors and embryonic development of porcine oocytes.
After 52 h of IVM, the phospho-p44/42 MAPK level was high in comparison with the total MAPK level (Figs. 1A and 1B). Our previous study demonstrated that 10 μM rapamycin increases blastocyst quality by affecting developmental rate and total cell number and reducing mitochondrial distribution, apoptosis, autophagy and ROS activity that regulates mTOR signaling [26]. The maternal genes expression levels of BMP15, GDF9, and MOS was higher in 0 R and 10 R oocytes than in control oocytes (Fig. 2A). Confirming the effect of rapamycin, levels of Cdc2 p34 and phospho-p44/42 MAPK were higher in 10 R oocytes than in control and 0 R oocytes (Figs. 2B, 2C, and 2D). MAPK and MPF are critical for meiotic molecular maturation of oocytes. Protein phosphorylation and dephosphorylation are essential for the meiotic cell cycle of oocytes. MPF and MAPK, which are integral components of the key regulatory pathways involved in activation of extracellular signal-regulated kinase 1 and 2, are serine/threonine kinases that are phosphorylated and subsequent activated by MAPK kinase [32,33]. MPF consists of the catalytic subunit p34/Cdc2, which possesses serine/threonine kinase activity, and the regulatory subunit cyclin B. Activation of MPF is regulated by various mechanisms, including binding of cyclin B to Cdc2, phosphorylation of threonine 161, and dephosphorylation of tyrosine 15 and threonine 14 [34]. MAPK activity usually peaks in porcine oocytes during IVM for 42–44 h. Many researchers have extended the IVM duration to improve oocyte maturity due to supply of insufficient maturation factors. The developmental rate reportedly increases when the IVM duration is extended (48–72 h). Specifically, the developmental rate is increased at 56 h due to maintenance of high MPF expression, but cytoplasmic senescence at 72 h decreases the developmental rate and suppresses MPF expression [35]. Another study confirmed that the developmental rate is improved by extending the IVM duration from 44 h, which is normally used. In the extended culture duration of 24 to 52 h, the cleavage rate of porcine oocytes was highest at 48 h as determined through the number of cumulus cell layers. [36]. Although that study did not reveal whether this approach affects a molecular factor, it showed that long-term culture is more effective for oocyte maturation [36]. To induce resumption of meiosis, porcine COCs at the pre-IVM stage were preincubated for 12 h [17]. Extension of the total IVM duration to 52 h increases the development rate of IVF embryos at the blastocyst stage [17]. Excessive prolongation of oocyte maturation leads to aging, and reduced MAPK activity decreases the quality of oocytes [37,38]. The IVM duration of poor-quality oocytes was previously extended to 52 h, and treatment with the antioxidant melatonin improved the IVM rate and expression levels of maturation factors [15]. We previously showed that treatment of aged oocytes with rapamycin increases phosphorylated p44/42 MAPK activity and mRNA levels of maternal genes compared with untreated aged oocytes [26]. Rapamycin activates MPF and MAPK, which reduces oocyte activation susceptibility via inhibition of protein kinase A [39]. Therefore, we suggest that rapamycin treatment facilitates molecular maturation by preserving the ooplasm of MII oocytes, leading to enhanced transcription of maternal genes. This study indicates that extension of the IVM duration with rapamycin treatment, which maintains expression of molecular maturation factors, does not negatively affect porcine oocytes.
ROS function are crucial signaling molecules in diverse physiological processes, including resumption of the meiotic cell cycle, and contribute to pathological processes such as apoptosis and senescence [40,41]. ROS were proposed to participate in oocyte meiotic arrest [42–44]. Oxidative stress perturbs bovine embryonic development after fertilization [45]. Furthermore, a change in the redox status of human oocytes during in vitro culture is related to an increased occurrence of apoptosis in gametes [46]. Porcine gametes can incur DNA damage and undergo apoptosis during IVM [47–49]. Treatment with rapamycin effectively reduces intracellular ROS levels and improves mitochondrial localization [26]. Extension of the IVM duration with rapamycin treatment demonstrated a significant reduction of levels of ROS (Figs. 3A and 3B). This finding suggests that this approach improves in vitro oocyte culture and maintains oocyte health.
We investigated the influence of extension of the IVM duration with rapamycin treatment on porcine embryonic development. The percentage of blastocyst formation at day 6 and total cell number per blastocyst were increased when the IVM duration was extended with rapamycin treatment (Figs. 4A, 4B, and 4C). Gene expression is a major contributor to embryonic development, and any disruption in gene expression during culture of embryos in vitro can potentially hinder embryo production [50,51]. We examined mRNA levels of POU5F1, SOX2, NANOG, and CDX2 in the 0 R, 10 R, and control groups. Transcription factor genes such as these play essential roles in early development and are indispensable for proliferation of undifferentiated embryonic stem (ES) cells in culture. mRNA and protein expression of POU5F1 has been detected in various cellular components, including blastomeres of preimplantation embryos, the ICM of blastocysts, epiblasts, primordial germ cells, and the majority of germ cells [52-54]. SOX2 and NANOG form interactions with POU5F1 to regulate the transcriptional hierarchy that specifies ES cell identity [55-57]. The transcription factors POU5F1 and SOX2 are expressed in both the ICM and trophectoderm (TE) of porcine blastocysts. CDX2 expression is essential for TE formation [58]. POU5F1 and NANOG exhibited significant upregulated (p < 0.05) in 10 R blastocysts (Fig. 4D). However, CDX2 expression was not significantly affected. These data show that extension of the IVM duration with rapamycin treatment upregulates specific transcription factors related to the ICM, including POU5F1 and NANOG, but does not affect expression of the TE-related factor CDX2 in blastocysts. Taken together, these observations demonstrate that extension of the IVM duration with rapamycin treatment significantly influences porcine embryonic development in vitro.
We explored the effect of extension of the IVM duration with rapamycin treatment on apoptotic cell death in porcine embryos. DNA fragmentation was significantly decreased in 10 R blastocysts extended duration of IVM by rapamycin treatment (Fig. 5B). We also examined the apoptosis expression levels of BCL2L1, FAS, and CASP3 (Fig. 5C). Although the pro-apoptotic expression levels of FAS and CASP3 was not different in all groups, the anti-apoptotic expression level of BCL2L1 in blastocysts was increased in the 10 R group. BCL2L1 encodes the protein BCL-xL, an anti-apoptotic protein [59]. BCL-xL prevents cell death by inhibiting cytochrome c release from mitochondria, a key step in the cell death pathway [60,61]. These observations suggest that extension of the IVM duration with rapamycin treatment facilitates embryonic development by suppressing apoptosis at the molecular level during preimplantation stages.
Our results indicate that extension of the IVM duration with rapamycin treatment enhances molecular maturation of porcine oocytes by repressing ROS production and improves porcine embryonic development. This study demonstrates the combination of extension of the IVM duration and treatment with rapamycin enhances maturation of porcine oocytes. These results should be further applied to assisted reproductive technology to produce high-quality embryos.